0% encontró este documento útil (0 votos)
62 vistas16 páginas2.3 Orgánulos Membranosos
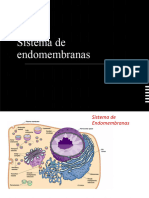
Este documento describe los diferentes tipos de orgánulos membranosos encontrados en las células eucariotas, incluyendo el retículo endoplasmático, el complejo de Golgi, lisosomas, mitocondrias y cloroplastos. Explica sus estructuras, funciones y roles en la síntesis de proteínas, metabolismo y digestión celular.
Cargado por
sofiamateo1808Derechos de autor
© © All Rights Reserved
Nos tomamos en serio los derechos de los contenidos. Si sospechas que se trata de tu contenido, reclámalo aquí.
Formatos disponibles
Descarga como PDF, TXT o lee en línea desde Scribd
0% encontró este documento útil (0 votos)
62 vistas16 páginas2.3 Orgánulos Membranosos
Este documento describe los diferentes tipos de orgánulos membranosos encontrados en las células eucariotas, incluyendo el retículo endoplasmático, el complejo de Golgi, lisosomas, mitocondrias y cloroplastos. Explica sus estructuras, funciones y roles en la síntesis de proteínas, metabolismo y digestión celular.
Cargado por
sofiamateo1808Derechos de autor
© © All Rights Reserved
Nos tomamos en serio los derechos de los contenidos. Si sospechas que se trata de tu contenido, reclámalo aquí.
Formatos disponibles
Descarga como PDF, TXT o lee en línea desde Scribd