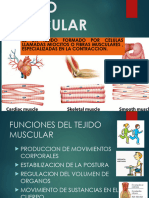
También podría gustarte
- Generalidades de Osteologia 2020Documento21 páginasGeneralidades de Osteologia 2020Anahi Rios100% (1)
- Cuestionarios Del Libro Ingeniería Industrial: Métodos, Estándares y Diseño Del TrabajoDocumento19 páginasCuestionarios Del Libro Ingeniería Industrial: Métodos, Estándares y Diseño Del TrabajoErica Rodriguez100% (2)
- Sistema óseo: Estructura, funciones y tipos de huesosDocumento6 páginasSistema óseo: Estructura, funciones y tipos de huesosEntrenador Isab100% (1)
- Sistema Óseo MorfologiaDocumento40 páginasSistema Óseo MorfologiaTomas ValdesAún no hay calificaciones
- OntogenesisDocumento12 páginasOntogenesisLorena VillanuevaAún no hay calificaciones
- Jorge Cancino Fisiología Del EjercicioDocumento34 páginasJorge Cancino Fisiología Del EjercicioKaren Torres PantojaAún no hay calificaciones
- Sistema Oseo Grupo 3Documento17 páginasSistema Oseo Grupo 3Mynor RodriguezAún no hay calificaciones
- Preguntas Repaso de FisiologiaDocumento12 páginasPreguntas Repaso de Fisiologiamindthegap2009100% (8)
- Taller de CarnicosDocumento17 páginasTaller de Carnicoserika reinaAún no hay calificaciones
- Como Tratar Y Prevenir El Dolor De Espalda: Descubre los factores que pueden ocasionarle dolores de espalda a una persona y cuáles son los posibles tratamientos para combatir este malDe EverandComo Tratar Y Prevenir El Dolor De Espalda: Descubre los factores que pueden ocasionarle dolores de espalda a una persona y cuáles son los posibles tratamientos para combatir este malCalificación: 5 de 5 estrellas5/5 (1)
- Tejido óseoDocumento3 páginasTejido óseoDubhé PeredaAún no hay calificaciones
- Aparato LocomotorDocumento3 páginasAparato LocomotorLAURA JINETH ACEVEDO CUERVOAún no hay calificaciones
- Fisiología del aparato locomotor: Células óseas y mecanismos de formación óseaDocumento31 páginasFisiología del aparato locomotor: Células óseas y mecanismos de formación óseaÁngel Javier Franco dominguezAún no hay calificaciones
- SISTEMA ESQUELÉTICO 4to Santa MariaDocumento6 páginasSISTEMA ESQUELÉTICO 4to Santa MariaFranz Lazcano TarquiAún no hay calificaciones
- República Boliv-WPS OfficeDocumento18 páginasRepública Boliv-WPS OfficeAlejandro PerezAún no hay calificaciones
- Traumatología ApuntesDocumento17 páginasTraumatología ApuntesMireya AnielaAún no hay calificaciones
- OSTEOLOGIADocumento28 páginasOSTEOLOGIACH JesúsAún no hay calificaciones
- FISIOLOGIA OSEADocumento4 páginasFISIOLOGIA OSEAgrethelcamila0615Aún no hay calificaciones
- Módulo 3: Huesos, músculos y articulacionesDocumento31 páginasMódulo 3: Huesos, músculos y articulacionesDepartamento AsistenciaAún no hay calificaciones
- OsteologiaDocumento7 páginasOsteologiaDavid PompaAún no hay calificaciones
- HuesoDocumento7 páginasHuesoMarco Vinicio Avendaño AlvarezAún no hay calificaciones
- Tejido óseo: estructura, funciones y tiposDocumento4 páginasTejido óseo: estructura, funciones y tiposRoberto RojasAún no hay calificaciones
- Revista Digital- Zona PelúcidaDocumento8 páginasRevista Digital- Zona PelúcidaTania ZaldivarAún no hay calificaciones
- Huesos: Fundamentos de Anatomía para Médicos: Estructura y Función Del HuesoDocumento15 páginasHuesos: Fundamentos de Anatomía para Médicos: Estructura y Función Del HuesoYANDIRA JANET SOSA IBAÑEZAún no hay calificaciones
- Practico 6B Biologia (Sistema Esqueletico)Documento15 páginasPractico 6B Biologia (Sistema Esqueletico)sofiaAún no hay calificaciones
- HuesosDocumento9 páginasHuesosMarck Anthony Ti EmAún no hay calificaciones
- Fisiologia Del HuesoDocumento7 páginasFisiologia Del HuesoJose012345Aún no hay calificaciones
- Sistema esquelético: estructura, funciones y formación de los huesosDocumento25 páginasSistema esquelético: estructura, funciones y formación de los huesosLeslie Johanna Buñay MazoAún no hay calificaciones
- Curso preparación física artes marcialesDocumento28 páginasCurso preparación física artes marcialesJavier BernalAún no hay calificaciones
- Tejido ÓseoDocumento94 páginasTejido ÓseoKarla M PerezAún no hay calificaciones
- Sistema Óseo y Su Relación Con El Aparato MuscularDocumento13 páginasSistema Óseo y Su Relación Con El Aparato MuscularYair MeraAún no hay calificaciones
- Estructura de Los HuesosDocumento9 páginasEstructura de Los HuesosMarco Velepucha100% (1)
- Sistema OseoDocumento20 páginasSistema OseoAnonymous g3Mz9CygCRAún no hay calificaciones
- Estructura de Los HuesosDocumento5 páginasEstructura de Los HuesosSarahi gonzalezAún no hay calificaciones
- Clase 3-Sistema Osteo-Artro-MuscularDocumento32 páginasClase 3-Sistema Osteo-Artro-MuscularKaren MaguidAún no hay calificaciones
- Generalidades de Los HuesosDocumento27 páginasGeneralidades de Los HuesosKarlos Albertoo Lauriano GutierrezAún no hay calificaciones
- Tejido OseoDocumento65 páginasTejido OseoEricka Danuskka SandovalAún no hay calificaciones
- Formación y estructura del tejido óseoDocumento6 páginasFormación y estructura del tejido óseoNita EscalonaAún no hay calificaciones
- Tejido ÓseoDocumento7 páginasTejido ÓseoSofía Cueva Cruz (Sofi)Aún no hay calificaciones
- Sistema OseoDocumento24 páginasSistema Oseotati4Aún no hay calificaciones
- Celulas Que Componen El Tejido OseoDocumento10 páginasCelulas Que Componen El Tejido OseoLisbeth VeintimilllaAún no hay calificaciones
- Trabajo de Los HuesosDocumento10 páginasTrabajo de Los HuesosNeimaryAún no hay calificaciones
- Informacion Del Sistema OseoDocumento2 páginasInformacion Del Sistema OseoLuis CarlosAún no hay calificaciones
- Los Huesos HumanosDocumento7 páginasLos Huesos HumanosMilagrosAún no hay calificaciones
- Tejido ÓseoDocumento6 páginasTejido Óseo202300183eAún no hay calificaciones
- Hueso Artificial Liofilizado Finall PDFDocumento53 páginasHueso Artificial Liofilizado Finall PDFJadiel QuispeAún no hay calificaciones
- Sistema OseoDocumento14 páginasSistema OseoCyntia CitlalliAún no hay calificaciones
- Tejido Conectivo (Histologia)Documento118 páginasTejido Conectivo (Histologia)GianninaPachecoAún no hay calificaciones
- Osteologia_GeneralidadesDocumento7 páginasOsteologia_GeneralidadesANARIA MEGOAún no hay calificaciones
- Sistema EsqueléticoDocumento83 páginasSistema Esqueléticonicolealbaortiz2003Aún no hay calificaciones
- Oseo y CartilaginosoDocumento7 páginasOseo y Cartilaginosoavedelterror100% (1)
- Sistema Esquelético 1Documento33 páginasSistema Esquelético 1alejandroAún no hay calificaciones
- Esqueleto axial y apendicular: estructura y funciones del sistema esqueléticoDocumento29 páginasEsqueleto axial y apendicular: estructura y funciones del sistema esqueléticoAlexandra CasariegoAún no hay calificaciones
- TEJIDO OSEO - Histologia IDocumento11 páginasTEJIDO OSEO - Histologia Iv52mhwg5x2Aún no hay calificaciones
- Cartilago y HuesoDocumento50 páginasCartilago y HuesoPablo Israel AverangaAún no hay calificaciones
- SISTEMA OSEO, Técnico en RadiologíaDocumento11 páginasSISTEMA OSEO, Técnico en RadiologíaPam OAún no hay calificaciones
- TEJIDO OSEO SeminarioDocumento41 páginasTEJIDO OSEO SeminarioDiego Salvador Regalado CastañedaAún no hay calificaciones
- Los Huesos y El EsqueletoDocumento45 páginasLos Huesos y El Esqueletow2s96r2tq4Aún no hay calificaciones
- CarreraDocumento5 páginasCarreraMary Isabel Hernández M.Aún no hay calificaciones
- Nuevo Documento de Microsoft WordDocumento11 páginasNuevo Documento de Microsoft WordPitter Garcia ColqueAún no hay calificaciones
- Modulo 11Documento9 páginasModulo 11vagosAún no hay calificaciones
- Comparto 'Bio 308-Módulo 3 Biología Del DesarrolloDocumento7 páginasComparto 'Bio 308-Módulo 3 Biología Del DesarrolloJavier HidalgoAún no hay calificaciones
- Bio 308-Guía de Lab.-Marta HigueraDocumento9 páginasBio 308-Guía de Lab.-Marta HigueraJavier HidalgoAún no hay calificaciones
- Lab. Preservación de Artrópodos y VertebradosDocumento24 páginasLab. Preservación de Artrópodos y VertebradosJavier HidalgoAún no hay calificaciones
- Congregaciones en PanamáDocumento14 páginasCongregaciones en PanamáJavier HidalgoAún no hay calificaciones
- Comparto 'Bio 308-Módulo 3 Biología Del DesarrolloDocumento7 páginasComparto 'Bio 308-Módulo 3 Biología Del DesarrolloJavier HidalgoAún no hay calificaciones
- Comparto 'Resumen de Aves (1) ' Con UstedDocumento13 páginasComparto 'Resumen de Aves (1) ' Con UstedJavier HidalgoAún no hay calificaciones
- Tabla Comparativa de Contraccion MusculoesqueleticolisoycardiacoDocumento7 páginasTabla Comparativa de Contraccion MusculoesqueleticolisoycardiacoKaren GomezAún no hay calificaciones
- Celulas Contractiles Kin-1Documento46 páginasCelulas Contractiles Kin-1Francisca Rubio LeivaAún no hay calificaciones
- Componentes de La CarneDocumento40 páginasComponentes de La CarneAnla LauraAún no hay calificaciones
- HistologíaDocumento89 páginasHistologíaAlfonso Martin SanchezAún no hay calificaciones
- Bases de Histología Médica PDFDocumento41 páginasBases de Histología Médica PDFCésar Colín50% (4)
- 3 Sistema MuscularDocumento47 páginas3 Sistema MuscularLuck ScordAún no hay calificaciones
- Fisiología de La Erección MasculinaDocumento2 páginasFisiología de La Erección MasculinaAndrés Felipe GonzálezAún no hay calificaciones
- Tejido MuscularDocumento12 páginasTejido Musculareuclydes BravoAún no hay calificaciones
- Histología - MúsculosDocumento4 páginasHistología - MúsculosIgnacio N. AlmazánAún no hay calificaciones
- Musculo 1Documento24 páginasMusculo 1Rafael PazAún no hay calificaciones
- Estructura y Función Del Musculo EsqueléticoDocumento7 páginasEstructura y Función Del Musculo EsqueléticoPiero MuñozAún no hay calificaciones
- Informe MuscularDocumento17 páginasInforme MuscularAlejandro AlvaradoAún no hay calificaciones
- Tejido Muscular HistologíaDocumento8 páginasTejido Muscular HistologíaAndrea Arriaga LimónAún no hay calificaciones
- El Papel de La Mecánica en Los Sistemas Biológicos y BioinspiradosDocumento10 páginasEl Papel de La Mecánica en Los Sistemas Biológicos y BioinspiradosSantiago AstudilloAún no hay calificaciones
- Cuáles Son Las Proteínas Presentes en La LecheDocumento12 páginasCuáles Son Las Proteínas Presentes en La LechePaulina Zamudio SarquiAún no hay calificaciones
- Fisiología de La Contracción UterinaDocumento18 páginasFisiología de La Contracción UterinaKelly MarcanoAún no hay calificaciones
- Contenido de Sistema Muscular2Documento8 páginasContenido de Sistema Muscular2Roland M ReyesAún no hay calificaciones
- 40 Tejido Muscular (Parte I y II)Documento50 páginas40 Tejido Muscular (Parte I y II)jpzk59cxprAún no hay calificaciones
- Clase4 Biomecanica IDocumento109 páginasClase4 Biomecanica IDANIEL ALEJANDROAún no hay calificaciones
- Citoesqueleto DiapositivasDocumento13 páginasCitoesqueleto DiapositivasDanii Escobar SantrichAún no hay calificaciones
- Prototipo ADocumento5 páginasPrototipo ARichard GarcíaAún no hay calificaciones
- Tejido muscular esquelético: estructura y funciónDocumento48 páginasTejido muscular esquelético: estructura y funciónMarcos DíasAún no hay calificaciones
- Músculo LisoDocumento13 páginasMúsculo LisoLaura Lizeth OspinaAún no hay calificaciones
- Características del tejido muscular (carne) y su estructuraDocumento28 páginasCaracterísticas del tejido muscular (carne) y su estructuraDiana Muñoz CisnerosAún no hay calificaciones
- Citoesquelto Resumen Cap12Documento9 páginasCitoesquelto Resumen Cap12claudiaAún no hay calificaciones
- Karol Resumen Clase 13Documento3 páginasKarol Resumen Clase 13Moises Castillo GAún no hay calificaciones