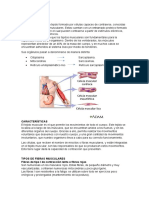
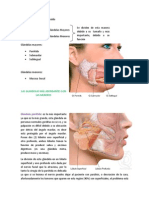
También podría gustarte
- Tejido Muscular Histologia 2019Documento4 páginasTejido Muscular Histologia 2019Ricardo SalasAún no hay calificaciones
- TEJIDO MUSCULAR Dra MaldonadoDocumento8 páginasTEJIDO MUSCULAR Dra MaldonadoLoismer LinaresAún no hay calificaciones
- Tejido Muscular Según La Evolución de Los AnimalesDocumento12 páginasTejido Muscular Según La Evolución de Los AnimalesInes Yulisa oviedo zuñigaAún no hay calificaciones
- Tejido MuscularDocumento11 páginasTejido MuscularGraciela Romero PalacioAún no hay calificaciones
- Tejido MuscularDocumento12 páginasTejido MuscularGerardo AcuñaAún no hay calificaciones
- Tarea: Histología Muscular: Sistema OsteomioarticularDocumento5 páginasTarea: Histología Muscular: Sistema OsteomioarticularHaraviel RiveraAún no hay calificaciones
- Copia de Tejido MuscularDocumento4 páginasCopia de Tejido MuscularEvelin Muñoz FretesAún no hay calificaciones
- Tejido Muscular Cardíaco y LisoDocumento7 páginasTejido Muscular Cardíaco y LisotomasAún no hay calificaciones
- Tejido muscular: estructura y tiposDocumento28 páginasTejido muscular: estructura y tiposCHRISTIAN ALBERTO AVILA TORRESAún no hay calificaciones
- Tejido MuscularDocumento12 páginasTejido MuscularMaria Virginia Alvarado LopezAún no hay calificaciones
- Tejido MuscularDocumento5 páginasTejido MuscularCris FernandezAún no hay calificaciones
- Tejido muscular: clasificación y funcionesDocumento8 páginasTejido muscular: clasificación y funcionesDARWIN RUDY CAMPERO GUZMÁNAún no hay calificaciones
- Sistemas musculares del cuerpo humanoDocumento7 páginasSistemas musculares del cuerpo humanoslinAún no hay calificaciones
- Tipos de Tejidos MuscularesDocumento4 páginasTipos de Tejidos Muscularesivanna5chalcobAún no hay calificaciones
- Clase 5Documento23 páginasClase 5Biol. Daniela López EscobarAún no hay calificaciones
- Cedva 14Documento21 páginasCedva 14Andrea Jimena Sosa CanoAún no hay calificaciones
- Tejido MuscularDocumento14 páginasTejido MuscularDay BravoAún no hay calificaciones
- Tejido MuscularDocumento6 páginasTejido MuscularMelody BarrosoAún no hay calificaciones
- Tejido MuscularDocumento2 páginasTejido MuscularSanAnimationsAún no hay calificaciones
- Cuáles Son Los Tipos de Tejido MuscularDocumento12 páginasCuáles Son Los Tipos de Tejido Muscularelfisico666Aún no hay calificaciones
- Tema 1. Sistema OsteomusculoarticularDocumento63 páginasTema 1. Sistema Osteomusculoarticularjulin400Aún no hay calificaciones
- Clase de Musculos.Documento85 páginasClase de Musculos.MARIAN8383Aún no hay calificaciones
- El Tejido MuscularDocumento3 páginasEl Tejido MuscularKAILER FIGUERAAún no hay calificaciones
- Musculo Estriado EsqueléticoDocumento3 páginasMusculo Estriado EsqueléticoCarolinaAún no hay calificaciones
- Representación Gráfica de Miocitos Según Clasificación Con DibujosDocumento3 páginasRepresentación Gráfica de Miocitos Según Clasificación Con DibujosAllison ZavalaAún no hay calificaciones
- Contraccion MuscularDocumento19 páginasContraccion MuscularAngela EscobarAún no hay calificaciones
- Tejido MuscularDocumento13 páginasTejido MusculardanielaAún no hay calificaciones
- Tejido Muscular HistologíaDocumento22 páginasTejido Muscular HistologíaLuiisPascaciioAún no hay calificaciones
- TEJIDOS MUSCULARES: CARACTERÍSTICAS Y TIPOSDocumento18 páginasTEJIDOS MUSCULARES: CARACTERÍSTICAS Y TIPOSAriana OroscoAún no hay calificaciones
- Tejido MuscularDocumento7 páginasTejido Musculartumaecom148Aún no hay calificaciones
- Tejido MuscularDocumento2 páginasTejido MuscularDaniela VásquezAún no hay calificaciones
- Tema 19 (Tejido Muscular)Documento37 páginasTema 19 (Tejido Muscular)Carlos Daniel Cárdenas SantosAún no hay calificaciones
- TEMA13 Tejidos AnimalDocumento4 páginasTEMA13 Tejidos AnimalClaudia Aldrey GolmarAún no hay calificaciones
- Músculos: Estructura, Tipos y FuncionesDocumento5 páginasMúsculos: Estructura, Tipos y FuncionesAlecksander TorresAún no hay calificaciones
- Tejido Muscular 198668 Downloable 1121332Documento5 páginasTejido Muscular 198668 Downloable 1121332Nayire Barboza HuatangareAún no hay calificaciones
- Semana 13Documento31 páginasSemana 13CRIS GONZÁLEZIAún no hay calificaciones
- Tejido Muscular Capitulo6Documento47 páginasTejido Muscular Capitulo6Raúl Ernesto Mendoza GranadosAún no hay calificaciones
- Brenda Tejido Muscular Por Dentro Del Titan ColosalDocumento6 páginasBrenda Tejido Muscular Por Dentro Del Titan ColosalJesus Abel Quiñonez ValenzuelaAún no hay calificaciones
- Tejido Nervioso y Tejido MuscularDocumento3 páginasTejido Nervioso y Tejido MuscularAriana León SalazarAún no hay calificaciones
- Tejido EpitelialDocumento18 páginasTejido EpitelialDavid Laiker MartiinezAún no hay calificaciones
- Tejido Muscular - Wikipedia, La Enciclopedia LibreDocumento9 páginasTejido Muscular - Wikipedia, La Enciclopedia LibreJesus CarreroAún no hay calificaciones
- Tjidos Del Cuuerpo Humano YsamarDocumento14 páginasTjidos Del Cuuerpo Humano Ysamarkike-alvinoAún no hay calificaciones
- Histologia Tejido Muscular ResumenDocumento5 páginasHistologia Tejido Muscular ResumenValentina Ortiz VarasAún no hay calificaciones
- Tejido MuscularDocumento54 páginasTejido MuscularAnonymous GgUTE2OAún no hay calificaciones
- Sistema muscular: clasificación, desarrollo y tipos de células muscularesDocumento14 páginasSistema muscular: clasificación, desarrollo y tipos de células muscularesCamilo HareaAún no hay calificaciones
- EnsayoTM LeonardoVelasquezDocumento7 páginasEnsayoTM LeonardoVelasquezleonardo velasquezAún no hay calificaciones
- Trabajo Tejido MuscularDocumento10 páginasTrabajo Tejido Muscularelismer gamez100% (1)
- Tejido MuscularDocumento9 páginasTejido MuscularOscar PerezAún no hay calificaciones
- Informe de Laboratorio 4. Grupo 6Documento15 páginasInforme de Laboratorio 4. Grupo 6christian vera0% (1)
- Anatomofisiologia Aplicada Clase 8Documento31 páginasAnatomofisiologia Aplicada Clase 8scarletAún no hay calificaciones
- Resumen Tejido MuscularDocumento4 páginasResumen Tejido MuscularANDREA CAMILA MARTINEZ GORDONAún no hay calificaciones
- Tejido Muscular ResumenDocumento21 páginasTejido Muscular ResumenEl KurkoAún no hay calificaciones
- Tejido MuscularDocumento5 páginasTejido MuscularCarolinaAún no hay calificaciones
- Tejido muscular: características y tiposDocumento6 páginasTejido muscular: características y tiposALEJANDRA VIVIANA RELICHE JIMENEZAún no hay calificaciones
- Tejido MuscularDocumento4 páginasTejido Muscularmaria tiguaAún no hay calificaciones
- Tejido MuscularDocumento9 páginasTejido MuscularDenysVillarrealAún no hay calificaciones
- Taler 1Documento7 páginasTaler 1GinaAún no hay calificaciones
- Fisio II (Guia 1)Documento12 páginasFisio II (Guia 1)manuel herradoraAún no hay calificaciones
- Tejido MuscularDocumento3 páginasTejido MuscularLuis Gustavo Rondon ChuecosAún no hay calificaciones
- Resumen Capítulos 2 y 3 - Aquiles BarrientosDocumento18 páginasResumen Capítulos 2 y 3 - Aquiles BarrientosAquiles Maxi BarrientosAún no hay calificaciones
- Quemaduras ElectricasDocumento42 páginasQuemaduras ElectricasJohn ChaucaAún no hay calificaciones
- Domador de CaballosDocumento11 páginasDomador de CaballosJulián HerbertAún no hay calificaciones
- Cancer de HipofaringeDocumento2 páginasCancer de HipofaringeAndyRamosLopezAún no hay calificaciones
- Itu AgudaDocumento28 páginasItu AgudaOffman VacaAún no hay calificaciones
- Sesionbibliografica JunioDocumento3 páginasSesionbibliografica JunioFilio OceranskyAún no hay calificaciones
- Guia OclusalDocumento3 páginasGuia OclusalFelix Fernando Caceres Morínigo33% (3)
- HomaopatiaDocumento19 páginasHomaopatiaAngiie PreyslerAún no hay calificaciones
- Tumores de OvarioDocumento62 páginasTumores de OvarioLetyDiaz100% (2)
- Posiciones QuirurgicasDocumento5 páginasPosiciones QuirurgicasAbel RodriguezAún no hay calificaciones
- Diagnóstico odontológico: exámen clínico y pruebas de sensibilidad pulparDocumento25 páginasDiagnóstico odontológico: exámen clínico y pruebas de sensibilidad pulparManuelito Guevara Vasquez100% (1)
- Analisis FacialDocumento4 páginasAnalisis FacialMaximiliano PerezAún no hay calificaciones
- Glandulas Salivales (Clase Desgrabada)Documento8 páginasGlandulas Salivales (Clase Desgrabada)Pablo RodriguezAún no hay calificaciones
- Operación dental: Aislamiento de campoDocumento113 páginasOperación dental: Aislamiento de campoRobertoChirinos100% (1)
- Dora La InvisibleDocumento2 páginasDora La InvisibleMishelleRodriguezBarcoAún no hay calificaciones
- Sistema DigestivoDocumento26 páginasSistema DigestivoSaúl MuroAún no hay calificaciones
- A. T. Mitchell - Cambiantes Oso 1 - Amar Al OsoDocumento166 páginasA. T. Mitchell - Cambiantes Oso 1 - Amar Al OsoMariana Barragan Gonzalez100% (1)
- Tema 7 - Nutrición en Animales I - Respiración y DigestiónDocumento38 páginasTema 7 - Nutrición en Animales I - Respiración y DigestiónMayra OrdoñezAún no hay calificaciones
- Ensayo KrakenDocumento5 páginasEnsayo KrakenЕлисо МарсианинAún no hay calificaciones
- Trichomonas TenaxDocumento10 páginasTrichomonas Tenaxanon_6099053530% (1)
- Guia Denticion TemporalDocumento4 páginasGuia Denticion TemporalAlejandro Vega100% (1)
- Flujograma Sistema Urinario PDFDocumento4 páginasFlujograma Sistema Urinario PDFPaula GamezAún no hay calificaciones
- Clase 1 Estomago IntestinoDocumento54 páginasClase 1 Estomago IntestinoKatherine Davila100% (1)
- Disfagia: causas, grados y clasificaciónDocumento2 páginasDisfagia: causas, grados y clasificaciónHugoSoto100% (1)
- T2 - Comunicación 1 - Equipo 2Documento3 páginasT2 - Comunicación 1 - Equipo 2Junior Alejandro Quispe EscalanteAún no hay calificaciones
- Anatomia Del CorazonDocumento5 páginasAnatomia Del CorazontifaniquirozAún no hay calificaciones
- El Violín Del Diablo, A Harry Potter Fanfic - FanFictionDocumento10 páginasEl Violín Del Diablo, A Harry Potter Fanfic - FanFictionnataliaar26Aún no hay calificaciones
- Taxonomía del Cóndor AndinoDocumento4 páginasTaxonomía del Cóndor AndinoFranklin SalinasAún no hay calificaciones
- Efectos Del Ruido en El Ser HumanoDocumento41 páginasEfectos Del Ruido en El Ser HumanoGonzalo Borgunder SoriaAún no hay calificaciones
- Leccion 2. EMBRIOLOGIA E HISTOLOGIA DENTAL PDFDocumento198 páginasLeccion 2. EMBRIOLOGIA E HISTOLOGIA DENTAL PDFEmmanuel CieloAún no hay calificaciones
- Psicología y trastornos de los niños en la edad evolutiva: Qué son y cómo funcionanDe EverandPsicología y trastornos de los niños en la edad evolutiva: Qué son y cómo funcionanCalificación: 5 de 5 estrellas5/5 (1)
- Neuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaDe EverandNeuroanatomía: Fundamentos de neuroanatomía estructural, funcional y clínicaCalificación: 4 de 5 estrellas4/5 (16)
- Notas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesDe EverandNotas de clase. Manual de farmacognosia: Análisis microscópico y fitoquímico, y usos de plantas medicinalesAún no hay calificaciones
- Dinámicas familiares a través de la vida de José: La túnica del PadreDe EverandDinámicas familiares a través de la vida de José: La túnica del PadreCalificación: 5 de 5 estrellas5/5 (2)
- Neurociencia para vencer la depresión: La esprial ascendenteDe EverandNeurociencia para vencer la depresión: La esprial ascendenteCalificación: 4.5 de 5 estrellas4.5/5 (10)
- Terapia cognitiva: Conceptos básicos y profundizaciónDe EverandTerapia cognitiva: Conceptos básicos y profundizaciónCalificación: 5 de 5 estrellas5/5 (1)
- Cómo conversar con cualquiera: Descubre cómo ser la persona más interesante del lugar y a destruir la ansiedad social permanentementeDe EverandCómo conversar con cualquiera: Descubre cómo ser la persona más interesante del lugar y a destruir la ansiedad social permanentementeCalificación: 4 de 5 estrellas4/5 (81)
- Terapia cognitiva del comportamiento: Descubre cómo despejar tu cerebro con la TCC. Aprender a superar el miedo y la ansiedad, la depresiónDe EverandTerapia cognitiva del comportamiento: Descubre cómo despejar tu cerebro con la TCC. Aprender a superar el miedo y la ansiedad, la depresiónCalificación: 5 de 5 estrellas5/5 (3)
- Educar la atención: Cómo entrenar esta habilidad en niños y adultosDe EverandEducar la atención: Cómo entrenar esta habilidad en niños y adultosCalificación: 4.5 de 5 estrellas4.5/5 (10)
- Mente Organizada Cómo Pensar con Claridad y Hacer Todas Las Decisiones de Vida CorrectasDe EverandMente Organizada Cómo Pensar con Claridad y Hacer Todas Las Decisiones de Vida CorrectasCalificación: 4 de 5 estrellas4/5 (11)
- Neuropsicología: Los fundamentos de la materiaDe EverandNeuropsicología: Los fundamentos de la materiaCalificación: 5 de 5 estrellas5/5 (1)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- DMT: La molécula del espíritu: Las revolucionarias investigaciones de un médico sobre la biología de las experiencias místicas y cercanas a la muerteDe EverandDMT: La molécula del espíritu: Las revolucionarias investigaciones de un médico sobre la biología de las experiencias místicas y cercanas a la muerteCalificación: 4 de 5 estrellas4/5 (135)
- Agilidad emocional: Rompe tus bloqueos, abraza el cambio y triunfa en el trabajo y en la vdaDe EverandAgilidad emocional: Rompe tus bloqueos, abraza el cambio y triunfa en el trabajo y en la vdaCalificación: 4.5 de 5 estrellas4.5/5 (34)
- Psicópatas seriales: Un recorrido por su oscura e inquietante naturalezaDe EverandPsicópatas seriales: Un recorrido por su oscura e inquietante naturalezaCalificación: 4 de 5 estrellas4/5 (3)
- Mindfulness: Cómo hacer menos y ser más conscientes puede ayudarnos a vivir mejor en este mundo estresadoDe EverandMindfulness: Cómo hacer menos y ser más conscientes puede ayudarnos a vivir mejor en este mundo estresadoCalificación: 4.5 de 5 estrellas4.5/5 (9)
- Nuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosDe EverandNuestra mente nos engaña: Sesgos y errores cognitivos que todos cometemosCalificación: 3.5 de 5 estrellas3.5/5 (6)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanDe EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanCalificación: 4.5 de 5 estrellas4.5/5 (11)
- El cerebro del niño explicado a los padresDe EverandEl cerebro del niño explicado a los padresCalificación: 4.5 de 5 estrellas4.5/5 (147)
- Cómo estimular el cerebro del niño: 100 ejercicios para potenciar la concentración, la memoria y otras funciones ejecutivasDe EverandCómo estimular el cerebro del niño: 100 ejercicios para potenciar la concentración, la memoria y otras funciones ejecutivasCalificación: 4.5 de 5 estrellas4.5/5 (12)
- Autismo: guía para padres sobre el trastorno del espectro autista En españolDe EverandAutismo: guía para padres sobre el trastorno del espectro autista En españolCalificación: 4.5 de 5 estrellas4.5/5 (4)
- Autoestima: Cómo evaluar, mejorar y mantener una sana autoestimaDe EverandAutoestima: Cómo evaluar, mejorar y mantener una sana autoestimaCalificación: 4.5 de 5 estrellas4.5/5 (10)
- El arte del descanso: Descubre el método para dormir bien y descansar mejorDe EverandEl arte del descanso: Descubre el método para dormir bien y descansar mejorCalificación: 4.5 de 5 estrellas4.5/5 (4)
- Bioneuroemoción: Un método para el bienestar emocionalDe EverandBioneuroemoción: Un método para el bienestar emocionalCalificación: 5 de 5 estrellas5/5 (4)
- Manual de psicoterapia emocional sistémicaDe EverandManual de psicoterapia emocional sistémicaCalificación: 4.5 de 5 estrellas4.5/5 (7)
- Cerebros rotos: Pacientes asombrosos que me enseñaron a vivirDe EverandCerebros rotos: Pacientes asombrosos que me enseñaron a vivirCalificación: 5 de 5 estrellas5/5 (3)