También podría gustarte
- Cómo calcular el tamaño de una muestra en investigación archivísticaDocumento6 páginasCómo calcular el tamaño de una muestra en investigación archivísticaPablo Antonio Daza YalicoAún no hay calificaciones
- Anestesia en Niños PDFDocumento4 páginasAnestesia en Niños PDFMilena Andrić100% (1)
- Anestésicos Locales 2018-1Documento4 páginasAnestésicos Locales 2018-1Nahuel Olivo100% (1)
- ALOP Lineamientos para El Uso de Terapias Pulpares en Dientes Primarios Con Pulpas No VitalesDocumento25 páginasALOP Lineamientos para El Uso de Terapias Pulpares en Dientes Primarios Con Pulpas No VitalesHilda Ayline Araya MorgadoAún no hay calificaciones
- Medicación intraconducto menos deDocumento65 páginasMedicación intraconducto menos deisis gomezAún no hay calificaciones
- 2021 I OdontogénesisDocumento36 páginas2021 I OdontogénesisAngel LampreaAún no hay calificaciones
- MelysSa PrOpEdEuTiCa2Documento53 páginasMelysSa PrOpEdEuTiCa2api-3723571Aún no hay calificaciones
- Anomalías Dentales Del Desarrollo y Erupción 5Documento22 páginasAnomalías Dentales Del Desarrollo y Erupción 5The Music Shine100% (1)
- CementomaDocumento6 páginasCementomaAna Diaz100% (1)
- Clase 3. Clasificación de Las Técnicas RadiográficasDocumento43 páginasClase 3. Clasificación de Las Técnicas RadiográficasChristopherAún no hay calificaciones
- Tipos de PupilasDocumento9 páginasTipos de PupilasDaniela CarbonellAún no hay calificaciones
- Guerrero Milady Español 3Documento36 páginasGuerrero Milady Español 3Elsy Nathali Vargas Vargas100% (1)
- Informe #2 Generalidades de ATM y Oclusiòn Dental.Documento28 páginasInforme #2 Generalidades de ATM y Oclusiòn Dental.Johanna Jaramillo MorenoAún no hay calificaciones
- Genesis, Identificacion y Uso de Los Suelos de MexicoDocumento244 páginasGenesis, Identificacion y Uso de Los Suelos de MexicoJluis Ipn100% (1)
- Trabajo Final Grupo 2Documento25 páginasTrabajo Final Grupo 2Bryam RosarioAún no hay calificaciones
- La piel: estructura, funciones y cuidadosDocumento7 páginasLa piel: estructura, funciones y cuidadosMariela TapiaAún no hay calificaciones
- UD 3. 2. Anatomía Dentaria II. Caras DentalesDocumento25 páginasUD 3. 2. Anatomía Dentaria II. Caras DentalesElena Sanchez BlancoAún no hay calificaciones
- Acido y BaseDocumento63 páginasAcido y Basedarkwolf1003Aún no hay calificaciones
- Coronas Estéticas en OdonropediatriaDocumento18 páginasCoronas Estéticas en OdonropediatriaGaby Mareño0% (1)
- Las Enfermedades Infectocontagiosas en La Consulta OdontológicaDocumento7 páginasLas Enfermedades Infectocontagiosas en La Consulta OdontológicaValentina VecchiottiAún no hay calificaciones
- SólidosDocumento34 páginasSólidosCindy Ochoa MaytorenaAún no hay calificaciones
- Manual de Calidad 2016Documento23 páginasManual de Calidad 2016victorAún no hay calificaciones
- PancreasDocumento1 páginaPancreasXiadani MenezAún no hay calificaciones
- Modulo Ecologia y Medio AmbienteDocumento85 páginasModulo Ecologia y Medio AmbienteWilfer Montes León100% (1)
- Libro Integrales Resueltas PDFDocumento242 páginasLibro Integrales Resueltas PDFJordan Hernandez Vidal100% (2)
- Reglas de DerivaciónDocumento17 páginasReglas de DerivaciónLeidy Marcela Gòmez MeloAún no hay calificaciones
- Polimeros Usados en La Undustria AlimentariaDocumento8 páginasPolimeros Usados en La Undustria AlimentariaEdgar BautistaAún no hay calificaciones
- Articulaciones Del CraneoDocumento1 páginaArticulaciones Del CraneoManuel HermidaAún no hay calificaciones
- Ceras en OdontologíaDocumento4 páginasCeras en Odontologíaana193Aún no hay calificaciones
- Bioseguridad Salud OralDocumento25 páginasBioseguridad Salud OralCarolina SarmientoAún no hay calificaciones
- Portafolio de ServiciosDocumento5 páginasPortafolio de ServiciosAna HenaoAún no hay calificaciones
- Clase 3 MantenimientoDocumento55 páginasClase 3 MantenimientoVtr Rjs0% (1)
- Glucogenolisis no es inverso de gluconeogénesisDocumento2 páginasGlucogenolisis no es inverso de gluconeogénesisMeybol Reyes CortezAún no hay calificaciones
- Capacitacion EspermaticaDocumento11 páginasCapacitacion EspermaticaSharon MosquedaAún no hay calificaciones
- 4 ProteinasDocumento11 páginas4 ProteinascanelgggAún no hay calificaciones
- Sindrome Compartimental AgudoDocumento5 páginasSindrome Compartimental AgudoDanielAntonioValderramaMerejildoAún no hay calificaciones
- GLANDULAS SALIVALES (Autoguardado)Documento26 páginasGLANDULAS SALIVALES (Autoguardado)eliasAún no hay calificaciones
- Expopsiscion Quitosano - QuimicaDocumento12 páginasExpopsiscion Quitosano - QuimicaAlejo MateusAún no hay calificaciones
- Criterios de ToxicidadDocumento1 páginaCriterios de ToxicidadAlejandro VazquezAún no hay calificaciones
- Una Región de Transición Asegura La Continuidad Vascular Entre Raíz y TalloDocumento4 páginasUna Región de Transición Asegura La Continuidad Vascular Entre Raíz y TalloStacy RosarioAún no hay calificaciones
- Odontogénesis OkDocumento2 páginasOdontogénesis OkJose D. ColqueAún no hay calificaciones
- Diseño de SonrisaDocumento3 páginasDiseño de Sonrisacomercializadora maglAún no hay calificaciones
- Farmacología de Los AntimicrobianosDocumento52 páginasFarmacología de Los AntimicrobianosJavier Heredia GodoyAún no hay calificaciones
- Factor Migración EspermáticaDocumento5 páginasFactor Migración EspermáticaRosilara SilvaAún no hay calificaciones
- Esteres y AmidasDocumento10 páginasEsteres y AmidasCristina Guerrero100% (1)
- Provisionales dentales: Materiales y técnicasDocumento5 páginasProvisionales dentales: Materiales y técnicasgriffo16Aún no hay calificaciones
- Metodo Falsa PosicionDocumento10 páginasMetodo Falsa PosicionRolando WalleAún no hay calificaciones
- Administracion Vs MarketingDocumento7 páginasAdministracion Vs MarketingAlicia Baluarte InocenteAún no hay calificaciones
- Postes PrefabricadosDocumento10 páginasPostes PrefabricadosRodríguez Roberto IvánAún no hay calificaciones
- DEF Solidos de RevolucionDocumento26 páginasDEF Solidos de RevolucionSaul Maravilla100% (1)
- CavidadesDocumento33 páginasCavidadesBzaAún no hay calificaciones
- Practica N 5Documento7 páginasPractica N 5kate vilcapaza CharcaAún no hay calificaciones
- Tema 4 LípidosDocumento15 páginasTema 4 LípidosYouAún no hay calificaciones
- NOM-005-SSA3-2018, Req Min de Infraestructura y EquipamientoDocumento11 páginasNOM-005-SSA3-2018, Req Min de Infraestructura y EquipamientoRodrigo EspinosaAún no hay calificaciones
- ENZIMASDocumento5 páginasENZIMASSebastián Camargo ReaAún no hay calificaciones
- Inhibidores enzimáticosDocumento21 páginasInhibidores enzimáticosAxel Ortiz CabanillasAún no hay calificaciones
- CUESTIONARIODocumento11 páginasCUESTIONARIOClaudia PalmaAún no hay calificaciones
- Resumen sobre las enzimas como catalizadores biológicosDocumento10 páginasResumen sobre las enzimas como catalizadores biológicosEnrique OrtizAún no hay calificaciones
- Cap 2 ENZIMASDocumento26 páginasCap 2 ENZIMASGianella Xiomara Valverde GómezAún no hay calificaciones
- Tema 9 EnzimasDocumento8 páginasTema 9 EnzimasSandrymar FloresAún no hay calificaciones
- Formatos PF en DigitalDocumento216 páginasFormatos PF en DigitalCésar Pablo Moran OrtizAún no hay calificaciones
- Formatos PF en DigitalDocumento216 páginasFormatos PF en DigitalCésar Pablo Moran OrtizAún no hay calificaciones
- Cinematica FisicaDocumento3 páginasCinematica FisicaCésar Pablo Moran OrtizAún no hay calificaciones
- Guía para El Seguimiento y Evaluación de Políticas Nacionales y Planes para El SINAPLAN - CEPLANDocumento110 páginasGuía para El Seguimiento y Evaluación de Políticas Nacionales y Planes para El SINAPLAN - CEPLANMarcos CardenasAún no hay calificaciones
- Analisis Dimensional FisicaDocumento3 páginasAnalisis Dimensional FisicaCésar Pablo Moran OrtizAún no hay calificaciones
- Nomenclatura QuimicaDocumento37 páginasNomenclatura Quimicaluis fernando moreno gutierrezAún no hay calificaciones
- Tratado de Criminalistica Tomo I PDFDocumento370 páginasTratado de Criminalistica Tomo I PDFcarlaAún no hay calificaciones
- Fibra Bruta y Fibra Dietetica OficialDocumento20 páginasFibra Bruta y Fibra Dietetica OficialRenzo Rodrigo Vilca Encinas100% (1)
- Taller Cálculos Con Ecuaciones Cúbicas de Estado para PetróleoDocumento16 páginasTaller Cálculos Con Ecuaciones Cúbicas de Estado para PetróleoSantiagoVélezSAún no hay calificaciones
- Guia Mayo 2021 PDFDocumento25 páginasGuia Mayo 2021 PDFinkpresionpublicidadAún no hay calificaciones
- Lubricacion PlasticaDocumento63 páginasLubricacion Plasticaauxiliar mantenimientoAún no hay calificaciones
- 3) Sabado 3 y Domingo 4 de Febrero 2024Documento24 páginas3) Sabado 3 y Domingo 4 de Febrero 2024Hebert RogelAún no hay calificaciones
- Densidad Del Lodo Del PerforacionDocumento13 páginasDensidad Del Lodo Del PerforacionJonathan Sucno IngaAún no hay calificaciones
- Practica 13enprocesoDocumento6 páginasPractica 13enprocesoJose SantamariaAún no hay calificaciones
- Titulación de soluciones ácidas y básicasDocumento5 páginasTitulación de soluciones ácidas y básicasYeison HernandezAún no hay calificaciones
- Tarea4.Operaciones UnitariasDocumento45 páginasTarea4.Operaciones UnitariasCielo Calderon GarciaAún no hay calificaciones
- Soluciones Buffer, Tampón o Amortiguadoras PDFDocumento11 páginasSoluciones Buffer, Tampón o Amortiguadoras PDFJeri11167% (6)
- Prueba Ley de Los Gases 8 Basico.aDocumento6 páginasPrueba Ley de Los Gases 8 Basico.aSusette DittusAún no hay calificaciones
- Silabo 2024-I QUÍMICA PARA INGENIEROS CDocumento23 páginasSilabo 2024-I QUÍMICA PARA INGENIEROS CÆđŕïäņ SerquenAún no hay calificaciones
- Solidos Cristalinos Terminado Power PointDocumento14 páginasSolidos Cristalinos Terminado Power PointPERCY DAVID MAXIMILIANO PAUCARAún no hay calificaciones
- Ejercicios Soluciones QuimicasDocumento2 páginasEjercicios Soluciones Quimicasmguadalupe2Aún no hay calificaciones
- Extraccion de CaseinaDocumento14 páginasExtraccion de CaseinaMelissa Calderon ArrietaAún no hay calificaciones
- Clasificación de compuestos orgánicos en hidrocarburos e hidrocarburos sustituidosDocumento2 páginasClasificación de compuestos orgánicos en hidrocarburos e hidrocarburos sustituidosdavoodoo89% (9)
- Memoria Cálculo Estanque Vertical 339m3Documento27 páginasMemoria Cálculo Estanque Vertical 339m3cristian toroAún no hay calificaciones
- Aplicaciones AminasDocumento3 páginasAplicaciones AminasNattiCrizzAún no hay calificaciones
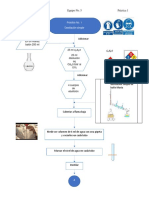
- Diagrama de Flujo P1Documento4 páginasDiagrama de Flujo P1VEGA FLORES Vianney IraisAún no hay calificaciones
- Cdm-Poc-06 Corte de Perfiles, Planchas, Tubos Con Equipo Oxiacetileno Rev.cDocumento13 páginasCdm-Poc-06 Corte de Perfiles, Planchas, Tubos Con Equipo Oxiacetileno Rev.cBryanAún no hay calificaciones
- Equilibrioproblemas 1Documento6 páginasEquilibrioproblemas 1AntonioGonzalezAún no hay calificaciones
- Caracteristicas de Las Familias de La Tabla PeriodicaDocumento5 páginasCaracteristicas de Las Familias de La Tabla PeriodicaGerman Soto Camacho72% (18)
- LAB-GYE-ME-403 Determinación de PH in SITU Revision 01Documento3 páginasLAB-GYE-ME-403 Determinación de PH in SITU Revision 01vanesaAún no hay calificaciones
- Química examen práctica preguntas reacciones átomosDocumento6 páginasQuímica examen práctica preguntas reacciones átomosfelipe M0% (1)
- Colorantes: compuestos que tiñen y mantienen colorDocumento11 páginasColorantes: compuestos que tiñen y mantienen colorRandyAún no hay calificaciones
- Agentes tóxicos: plomo, mercurio, etanol, tolueno y monóxido de carbonoDocumento15 páginasAgentes tóxicos: plomo, mercurio, etanol, tolueno y monóxido de carbonoKarol Vanessa MOSQUERA JARAMILLOAún no hay calificaciones
- Guia ComplementariaDocumento2 páginasGuia ComplementariaGabriela RegelAún no hay calificaciones
- Mario Rodriguez TesisDocumento69 páginasMario Rodriguez TesisDiego Chicaíza FinleyAún no hay calificaciones
- La revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaDe EverandLa revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaCalificación: 5 de 5 estrellas5/5 (200)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanDe EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Investigación de operaciones: Conceptos fundamentalesDe EverandInvestigación de operaciones: Conceptos fundamentalesCalificación: 4.5 de 5 estrellas4.5/5 (2)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDe EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceCalificación: 5 de 5 estrellas5/5 (8)
- Reconstrucción de dientes endodonciados: Pautas de actuación clínicaDe EverandReconstrucción de dientes endodonciados: Pautas de actuación clínicaCalificación: 5 de 5 estrellas5/5 (4)
- Psiconeuroinmunología para la práctica clínicaDe EverandPsiconeuroinmunología para la práctica clínicaCalificación: 5 de 5 estrellas5/5 (4)
- La invasión de la pseudociencia: Claves para orientarse en un mundo en donde casi todo es posibleDe EverandLa invasión de la pseudociencia: Claves para orientarse en un mundo en donde casi todo es posibleCalificación: 4.5 de 5 estrellas4.5/5 (15)
- Nunca fuimos modernos: Ensayos de antropología simétricaDe EverandNunca fuimos modernos: Ensayos de antropología simétricaAún no hay calificaciones
- Disciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreDe EverandDisciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreCalificación: 5 de 5 estrellas5/5 (3)
- Interpretación del ECG: Una Guía Práctica e Intuitiva para Aprender a Leer el ECG y Diagnosticar y Tratar ArritmiasDe EverandInterpretación del ECG: Una Guía Práctica e Intuitiva para Aprender a Leer el ECG y Diagnosticar y Tratar ArritmiasAún no hay calificaciones
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Genética general: Libro de textoDe EverandGenética general: Libro de textoCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Manual práctico de medicina tradicional china para cada día: Tés nutritivos y curativos, sencillos ejercicios de Tai-Chi y Qi-Gong, acupresión, meditación y… mucho másDe EverandManual práctico de medicina tradicional china para cada día: Tés nutritivos y curativos, sencillos ejercicios de Tai-Chi y Qi-Gong, acupresión, meditación y… mucho másCalificación: 5 de 5 estrellas5/5 (2)
- Anatomía & 100 estiramientos Esenciales (Color): Técnicas, beneficios, precauciones, consejos, tablas de series, dolenciasDe EverandAnatomía & 100 estiramientos Esenciales (Color): Técnicas, beneficios, precauciones, consejos, tablas de series, dolenciasCalificación: 4.5 de 5 estrellas4.5/5 (21)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaDe Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaCalificación: 4 de 5 estrellas4/5 (37)
- Estadística básica: Introducción a la estadística con RDe EverandEstadística básica: Introducción a la estadística con RCalificación: 5 de 5 estrellas5/5 (8)
- Fisiopatología de las enfermedades cardiovascularesDe EverandFisiopatología de las enfermedades cardiovascularesCalificación: 5 de 5 estrellas5/5 (1)
- El libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)De EverandEl libro conciso de la punción seca: Manual del terapeuta para las aplicaciones en los puntos gatillo miofasciales (Color)Calificación: 3 de 5 estrellas3/5 (2)
- En busca de la mente: El largo camino para comprender la vida mental (y lo que aún queda por descubrir)De EverandEn busca de la mente: El largo camino para comprender la vida mental (y lo que aún queda por descubrir)Calificación: 4.5 de 5 estrellas4.5/5 (3)