También podría gustarte
- Dinosauria1 6Documento8 páginasDinosauria1 6sinbuAún no hay calificaciones
- Dinosauria1 2Documento8 páginasDinosauria1 2sinbuAún no hay calificaciones
- Átomo 1 3Documento5 páginasÁtomo 1 3sinbuAún no hay calificaciones
- Dinosauria1 5Documento7 páginasDinosauria1 5sinbuAún no hay calificaciones
- Dinosauria1 3Documento8 páginasDinosauria1 3sinbuAún no hay calificaciones
- Dinosauria1 1Documento7 páginasDinosauria1 1sinbuAún no hay calificaciones
- La MoleculaDocumento7 páginasLa MoleculasinbuAún no hay calificaciones
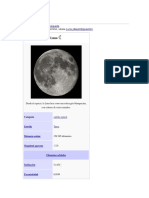
- La Luna FinalizaDocumento7 páginasLa Luna FinalizasinbuAún no hay calificaciones
- Monstruo MarinoDocumento3 páginasMonstruo Marinosinbu0% (1)
- Luna: satélite natural de la TierraDocumento15 páginasLuna: satélite natural de la TierrasinbuAún no hay calificaciones
- El Conquistador Del Fin Del MundoDocumento3 páginasEl Conquistador Del Fin Del MundosinbuAún no hay calificaciones
- Los PayasosDocumento6 páginasLos PayasossinbuAún no hay calificaciones
- Átomo 1 2Documento6 páginasÁtomo 1 2sinbuAún no hay calificaciones
- Átomo 1 1Documento7 páginasÁtomo 1 1sinbuAún no hay calificaciones
- Segunda Guerra Mundial (WW II) - Parte3Documento7 páginasSegunda Guerra Mundial (WW II) - Parte3sinbuAún no hay calificaciones
- La Luna SigueDocumento8 páginasLa Luna SiguesinbuAún no hay calificaciones
- (WW II) - Parte5Documento7 páginas(WW II) - Parte5sinbuAún no hay calificaciones
- Guerra Civil Española y ascenso del nazismoDocumento7 páginasGuerra Civil Española y ascenso del nazismosinbuAún no hay calificaciones
- La Historia de Mexico-Parte4Documento4 páginasLa Historia de Mexico-Parte4sinbuAún no hay calificaciones
- (WW II) - Parte6Documento8 páginas(WW II) - Parte6sinbuAún no hay calificaciones
- Segunda Guerra Mundial (WW II) - Parte1Documento7 páginasSegunda Guerra Mundial (WW II) - Parte1sinbuAún no hay calificaciones
- La Historia de Mexico-Parte2Documento8 páginasLa Historia de Mexico-Parte2sinbuAún no hay calificaciones
- La Historia de Mexico-Parte1Documento8 páginasLa Historia de Mexico-Parte1sinbuAún no hay calificaciones
- Revolución Mexicana PARTE 1Documento8 páginasRevolución Mexicana PARTE 1sinbuAún no hay calificaciones
- La Historia de Mexico-Parte1Documento8 páginasLa Historia de Mexico-Parte1sinbuAún no hay calificaciones
- Independencia de México PARTE 10Documento6 páginasIndependencia de México PARTE 10sinbuAún no hay calificaciones
- La Historia de Mexico-Parte3Documento8 páginasLa Historia de Mexico-Parte3sinbuAún no hay calificaciones
- Independencia de México PARTE 9Documento7 páginasIndependencia de México PARTE 9sinbuAún no hay calificaciones
- Independencia de México PARTE 8Documento8 páginasIndependencia de México PARTE 8sinbuAún no hay calificaciones
- Las ComillasDocumento3 páginasLas ComillasMael Sasa Jccm0% (1)
- Reforzamiento por tema común 6° grado semana 27Documento21 páginasReforzamiento por tema común 6° grado semana 27María HOAún no hay calificaciones
- Sistemas OperativosDocumento157 páginasSistemas OperativosANDRES BRAVO AGUILARAún no hay calificaciones
- Diseño de Un Plan de Marketing Acorde A Las Necesidades de Las Mí Pymes Del Sector Comercial de La Ciudad de Barranquilla y Su Área MetropolitanaDocumento78 páginasDiseño de Un Plan de Marketing Acorde A Las Necesidades de Las Mí Pymes Del Sector Comercial de La Ciudad de Barranquilla y Su Área MetropolitanaMoises AcuñaAún no hay calificaciones
- Chiavaneto Recursos HumanosDocumento11 páginasChiavaneto Recursos HumanosJhon RuizAún no hay calificaciones
- Contaminación océano metales fertilizantes impacto ecosistemaDocumento12 páginasContaminación océano metales fertilizantes impacto ecosistemaanizaAún no hay calificaciones
- 1-5° El CuentoDocumento3 páginas1-5° El CuentoIsmelda0% (1)
- Lab04 521230 2018 PDFDocumento10 páginasLab04 521230 2018 PDFAndres LAún no hay calificaciones
- Abdomen Agudo Dr. Andrés Volio C. Residente de Cirugía GeneralDocumento50 páginasAbdomen Agudo Dr. Andrés Volio C. Residente de Cirugía Generalpaola bendezu saraviaAún no hay calificaciones
- Asesoria de Impacto HogaramaDocumento7 páginasAsesoria de Impacto HogaramaJose GuzmanAún no hay calificaciones
- Derecho Internacional Del TrabajoDocumento6 páginasDerecho Internacional Del TrabajoAndre BlazquezAún no hay calificaciones
- Modulo Matematicas FinancierasDocumento166 páginasModulo Matematicas Financierassalazar_edwardAún no hay calificaciones
- Estatutos Internos para La Conformacion y El Funcionamiento Del CSSLDocumento10 páginasEstatutos Internos para La Conformacion y El Funcionamiento Del CSSLJohan Manuel Sanchez MorenoAún no hay calificaciones
- Tipos de Dominación Max WeberDocumento2 páginasTipos de Dominación Max WeberEhudd Romero67% (3)
- IND414 - Manual de Diseño Inst Dom AP en EDIF - 250520 PDFDocumento75 páginasIND414 - Manual de Diseño Inst Dom AP en EDIF - 250520 PDFalfonso murguiaAún no hay calificaciones
- Bases de Medicion Unitarias y Multimedicion EZMDocumento10 páginasBases de Medicion Unitarias y Multimedicion EZMe34219351Aún no hay calificaciones
- Caso Javier 20Documento3 páginasCaso Javier 20Giuli D'AngeloAún no hay calificaciones
- Cuaderno IICE 13 FINALDocumento117 páginasCuaderno IICE 13 FINALSDFSDJKFJKNAún no hay calificaciones
- QualiaDocumento2 páginasQualiaJulioAranedaAún no hay calificaciones
- El Divino Don Del Perdón PDFDocumento171 páginasEl Divino Don Del Perdón PDFDemian Gabriela Osorio Gallardo100% (11)
- Competencia Imperfecta, RobinsonDocumento5 páginasCompetencia Imperfecta, RobinsonPEDRO MANUEL LEMUS PARDOAún no hay calificaciones
- El Rey Azúcar y Otros Monarcas AgrícolasDocumento1 páginaEl Rey Azúcar y Otros Monarcas AgrícolasNeicerAún no hay calificaciones
- SueloDocumento18 páginasSueloSotilllo Yerbes UbencioAún no hay calificaciones
- Cuadernillo Pendientes 3 ESO2 Evaluacion 2014Documento13 páginasCuadernillo Pendientes 3 ESO2 Evaluacion 2014Lucia Borràs PruñonosaAún no hay calificaciones
- Posiciones AnatomicasDocumento3 páginasPosiciones AnatomicasVictoria RomeroAún no hay calificaciones
- Motores de combustión interna: prueba diagnósticaDocumento2 páginasMotores de combustión interna: prueba diagnósticaXavier Guaman AgudoAún no hay calificaciones
- Max Weber: El origen del espíritu capitalistaDocumento56 páginasMax Weber: El origen del espíritu capitalistaAna BonlluvanniAún no hay calificaciones
- Prácticas agronómicas I UNALMDocumento8 páginasPrácticas agronómicas I UNALMGustavo Alonzo Samame GoicocheaAún no hay calificaciones
- TESIS ACTUARIA (13) - 28 de Junio de 2017 Dulce Ivette Herrera HerreraDocumento132 páginasTESIS ACTUARIA (13) - 28 de Junio de 2017 Dulce Ivette Herrera Herrerajose zepedaAún no hay calificaciones
- Hábitos alimenticios del personal de enfermería durante su jornada laboralDocumento34 páginasHábitos alimenticios del personal de enfermería durante su jornada laboralFederico BeverAún no hay calificaciones