También podría gustarte
- Ensayo de LipidosDocumento13 páginasEnsayo de LipidosArturo Hernández Sandoval0% (2)
- Actividad Proteinas Grupo 16Documento11 páginasActividad Proteinas Grupo 16Dianaa Isabella FernandezAún no hay calificaciones
- Actividad Especial Bioquímica de ProteínaDocumento20 páginasActividad Especial Bioquímica de ProteínaDianaa Isabella FernandezAún no hay calificaciones
- Regulacion Del Bombeo CardiacoDocumento3 páginasRegulacion Del Bombeo CardiacoDianaa Isabella FernandezAún no hay calificaciones
- Electrodo de Membrana Líquida para CalcioDocumento2 páginasElectrodo de Membrana Líquida para CalcioDianaa Isabella Fernandez0% (1)
- Parcial 2Documento88 páginasParcial 2Dianaa Isabella FernandezAún no hay calificaciones
- Preguntas de Bioquimica 2Documento5 páginasPreguntas de Bioquimica 2Aguirre Carrillo Maria CelesteAún no hay calificaciones
- Cuestionario de Autoevaluación Del TP 4Documento11 páginasCuestionario de Autoevaluación Del TP 4iasmin26Aún no hay calificaciones
- Citoplasma Citosol Sin AudioDocumento20 páginasCitoplasma Citosol Sin AudioFlorencia CastilloAún no hay calificaciones
- Absorcion NutrientesDocumento19 páginasAbsorcion Nutrientesdeybi_torresAún no hay calificaciones
- BombasDocumento22 páginasBombasNynoshka CoveñasAún no hay calificaciones
- Mioglobina MonoDocumento6 páginasMioglobina MonoXimena Ticona Justo100% (1)
- Reacciones de TransaminaciónDocumento3 páginasReacciones de TransaminaciónJan Alexis Cedillo Alcantara100% (1)
- Estructura de Las ProteínasDocumento4 páginasEstructura de Las ProteínasThelma Flores ZapataAún no hay calificaciones
- Enzimas PDFDocumento53 páginasEnzimas PDFDiego TorresAún no hay calificaciones
- UntitledDocumento10 páginasUntitledBRYAN ALEXIS ORDOÑEZ RECINOSAún no hay calificaciones
- B-Arrestin Mediated Receptor Trafficking and Signal TraductionDocumento13 páginasB-Arrestin Mediated Receptor Trafficking and Signal TraductionSILVIA FERNANDA CASTILLO GOYENECHEAún no hay calificaciones
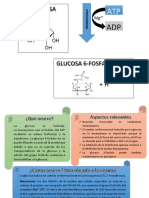
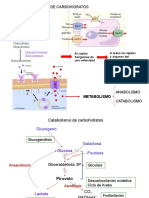
- Cuestionario Metabolismo CarbohidratosDocumento7 páginasCuestionario Metabolismo CarbohidratosfanyAún no hay calificaciones
- Comparación de Métodos Analíticos en El Laboratorio ClínicoDocumento5 páginasComparación de Métodos Analíticos en El Laboratorio Clínicoandrew sotoAún no hay calificaciones
- 3 QDP 6 Edlc ProteínasDocumento41 páginas3 QDP 6 Edlc ProteínasIsa Linares de la CalAún no hay calificaciones
- PT y aPTT - Ensayo de Primera Línea para Análisis de Coagulación - Hemostasia.Documento6 páginasPT y aPTT - Ensayo de Primera Línea para Análisis de Coagulación - Hemostasia.Juan Sin TierraAún no hay calificaciones
- Valores de ReferenciaDocumento1 páginaValores de ReferencianathyAún no hay calificaciones
- CAP 5 - Biologia CelularDocumento31 páginasCAP 5 - Biologia CelularClara RamirezAún no hay calificaciones
- Laboratorio VillaDocumento76 páginasLaboratorio VillaJimmy Silvestre Corpus AcostaAún no hay calificaciones
- Und - Planeador de Clase Grado Undécimo de Quìmica Segundo Semestre - 2021Documento3 páginasUnd - Planeador de Clase Grado Undécimo de Quìmica Segundo Semestre - 2021Freddy PeñareteAún no hay calificaciones
- Mapa Conceptual de Las BiomoleculasDocumento1 páginaMapa Conceptual de Las BiomoleculasLX EvmAún no hay calificaciones
- Aminoácidos y Proteína - Orgánica IIIDocumento62 páginasAminoácidos y Proteína - Orgánica IIILUIS CARLOS ROMERO ZAPATAAún no hay calificaciones
- 2.1.1 Glucólisis. - Ruta MetabólicaDocumento20 páginas2.1.1 Glucólisis. - Ruta MetabólicaANDREA CORDEROAún no hay calificaciones
- Clases 06 LÍPIDOSDocumento25 páginasClases 06 LÍPIDOSEVELIN LIZBETH VALLEJOS SANTA CRUZAún no hay calificaciones
- HOJA DE TRABAJO Perfil Lipídico 2Documento7 páginasHOJA DE TRABAJO Perfil Lipídico 2Chiara GonzalesAún no hay calificaciones
- Digestión de Carbohidratos, Proteínas y LípidosDocumento36 páginasDigestión de Carbohidratos, Proteínas y LípidosJose TapiaAún no hay calificaciones
- Fuerzas Implicadas en La Estabilidad de La Estructura de Las ProteínasDocumento2 páginasFuerzas Implicadas en La Estabilidad de La Estructura de Las ProteínasConstanza Coronado RiveraAún no hay calificaciones
- Unidad2-2 2022Documento30 páginasUnidad2-2 2022antonio reyesAún no hay calificaciones
- Oxidación de Ácidos Grasos - CetogénesisDocumento6 páginasOxidación de Ácidos Grasos - CetogénesisKarelys Brigitte Tandazo AtancuriAún no hay calificaciones
- Cadena Respiratoria y Fosforilación Oxidativa 2012Documento26 páginasCadena Respiratoria y Fosforilación Oxidativa 2012Hugo VillalbaAún no hay calificaciones