También podría gustarte
- Curso PRP Medicina EsteticaDocumento47 páginasCurso PRP Medicina EsteticaRafael ZambranoAún no hay calificaciones
- Especializaciones de MembranaDocumento6 páginasEspecializaciones de Membranadaniela1dcp92% (26)
- DX Labor y PartoDocumento50 páginasDX Labor y PartoJose M Hernandez100% (1)
- Solucion Examen Biologia Selectividad Andalucia Junio 2021 PDFDocumento7 páginasSolucion Examen Biologia Selectividad Andalucia Junio 2021 PDFRobAún no hay calificaciones
- Fase Luminosa Fot FichaDocumento2 páginasFase Luminosa Fot FichaProf. Víctor M. VitoriaAún no hay calificaciones
- TEMA 3 Nutrición y AlimentaciónDocumento4 páginasTEMA 3 Nutrición y AlimentaciónAlejandro FernandezAún no hay calificaciones
- Bloque 5 InmunologíaDocumento16 páginasBloque 5 InmunologíadlkjfvAún no hay calificaciones
- Biologia Programacio CCNN 1º y 3º 22 23 Documentos de GoogleDocumento129 páginasBiologia Programacio CCNN 1º y 3º 22 23 Documentos de GoogleEncarnaAún no hay calificaciones
- Una Nueva Suavidad-RolnikDocumento3 páginasUna Nueva Suavidad-RolnikJulia Broguet100% (1)
- Examen Resuelto PAU Biología 2018 Junio - Solución EJERCICIOS Y EXÁMENES RESUELTOSDocumento1 páginaExamen Resuelto PAU Biología 2018 Junio - Solución EJERCICIOS Y EXÁMENES RESUELTOSOlocauAún no hay calificaciones
- Examen Resuelto Biología Selectividad Junio 2022 AndalucíaDocumento8 páginasExamen Resuelto Biología Selectividad Junio 2022 AndalucíaSonia CalatayudAún no hay calificaciones
- TEMA 10 ReplicaciónDocumento6 páginasTEMA 10 ReplicaciónLAURA LARAAún no hay calificaciones
- Apuntes PDFDocumento43 páginasApuntes PDFJhonatan Torres CastroAún no hay calificaciones
- Informe Psicologico Del Test RorschachDocumento2 páginasInforme Psicologico Del Test Rorschachedson crisostomo100% (1)
- Dymind DM79XDocumento4 páginasDymind DM79XCasique OldemarAún no hay calificaciones
- Orientaciones Biología Selectividad 2021-2022Documento17 páginasOrientaciones Biología Selectividad 2021-2022Sophia El HarrakAún no hay calificaciones
- Biología Apuntes de La EBAU 2020Documento4 páginasBiología Apuntes de La EBAU 2020NereaAún no hay calificaciones
- Examen Ebau de Biología Junio 2022Documento4 páginasExamen Ebau de Biología Junio 2022a aaAún no hay calificaciones
- Recomendaciones y Orientaciones Biología MAYORES2021Documento125 páginasRecomendaciones y Orientaciones Biología MAYORES2021tonitot8Aún no hay calificaciones
- Apuntes de Selectividad, BiologíaDocumento4 páginasApuntes de Selectividad, BiologíaSebastián BurgosAún no hay calificaciones
- Examen Biologia Ebau Julio 21Documento4 páginasExamen Biologia Ebau Julio 21Alba Maria Cardona BethencourtAún no hay calificaciones
- Ficha de Estudio Lisosomas y PeroxisomasDocumento2 páginasFicha de Estudio Lisosomas y PeroxisomasProf. Víctor M. Vitoria100% (1)
- Ebau Microbiologia Soluciones 2020Documento16 páginasEbau Microbiologia Soluciones 2020Pedro MartínezAún no hay calificaciones
- Biologia Selectividad Examen Resuelto Canarias 4 WWW - Siglo21x.blogspotDocumento8 páginasBiologia Selectividad Examen Resuelto Canarias 4 WWW - Siglo21x.blogspotmonografiasxAún no hay calificaciones
- Exámenes Resueltos Biología 2 Bachilllerato UNED Modelo03 2015Documento3 páginasExámenes Resueltos Biología 2 Bachilllerato UNED Modelo03 2015Ivaj El HumanoAún no hay calificaciones
- Tema 1 (La Célula Procariota) PDFDocumento16 páginasTema 1 (La Célula Procariota) PDFNonde OneneAún no hay calificaciones
- 08 Membranas 2 Bach PDFDocumento9 páginas08 Membranas 2 Bach PDFPaco NavarroAún no hay calificaciones
- Nomenclatura Iupac Organica JaDocumento19 páginasNomenclatura Iupac Organica JaweikeAún no hay calificaciones
- BLOQUE II - Biologia 2 PEvAU Andalucia 201819Documento1 páginaBLOQUE II - Biologia 2 PEvAU Andalucia 201819Cristina Silva LópezAún no hay calificaciones
- Resumen Fisiologia CelularDocumento8 páginasResumen Fisiologia CelularMario Javier Ganoza Morales100% (1)
- Curriculum Bachillerato Biología LOMCE, Castilla y LeónDocumento17 páginasCurriculum Bachillerato Biología LOMCE, Castilla y LeónJoséLuisTorresGonzálezAún no hay calificaciones
- Quimica OrganicaDocumento14 páginasQuimica Organicadaniel andresAún no hay calificaciones
- Poríferos Cnidarios Platelmintos Nematodos 1bachDocumento16 páginasPoríferos Cnidarios Platelmintos Nematodos 1bachBiomlio Mijoy NimjiAún no hay calificaciones
- Biologia Selectividad Examen 5 Resuelto Castilla y Leon WWW - Siglo21x.blogspotDocumento6 páginasBiologia Selectividad Examen 5 Resuelto Castilla y Leon WWW - Siglo21x.blogspotmonografiasxAún no hay calificaciones
- MatematicasAplicadasCCSSII SolucionDocumento9 páginasMatematicasAplicadasCCSSII SolucioncangalfAún no hay calificaciones
- 03 Glucidos 2 BachDocumento8 páginas03 Glucidos 2 BachfsarrodAún no hay calificaciones
- Ejercicios Resueltos Tema 9 Fisica y Quimica 4º ESODocumento19 páginasEjercicios Resueltos Tema 9 Fisica y Quimica 4º ESOJose Antonio Coronado TorresAún no hay calificaciones
- Ebook en PDF BIOLOGIA PREGUNTAS DE SELECTIVIDAD ANDALUCIA 2001 2012Documento183 páginasEbook en PDF BIOLOGIA PREGUNTAS DE SELECTIVIDAD ANDALUCIA 2001 2012SHANYIN DANAE REYES GARCIAAún no hay calificaciones
- T-31 Replicación Del DNA, Transcripción, Código Genético Y TRADUCCIONDocumento27 páginasT-31 Replicación Del DNA, Transcripción, Código Genético Y TRADUCCIONYajaira SeguraAún no hay calificaciones
- Examen Matematicas II Ebau Junio 2022Documento2 páginasExamen Matematicas II Ebau Junio 2022Alejandro CedresAún no hay calificaciones
- Practicas HistologíaDocumento40 páginasPracticas HistologíaYulsAún no hay calificaciones
- Ejercicios de GenéticaDocumento32 páginasEjercicios de GenéticaSheila GarcíaAún no hay calificaciones
- Actividades Evau Bloque 1 - BiomoléculasDocumento7 páginasActividades Evau Bloque 1 - BiomoléculasAaronfuenla 15Aún no hay calificaciones
- Sol REPASO CURSO BIOLOGÍA 2011Documento25 páginasSol REPASO CURSO BIOLOGÍA 2011Prof. Víctor M. Vitoria100% (1)
- Cuader Bio 4 NewDocumento36 páginasCuader Bio 4 NewAlfredo Bautista HernándezAún no hay calificaciones
- Examen Biologia Selectividad 2017 Enunciado y SolucionDocumento8 páginasExamen Biologia Selectividad 2017 Enunciado y SolucionM MN0% (1)
- Acidos Nucleicos 17-18-170910104832 PDFDocumento17 páginasAcidos Nucleicos 17-18-170910104832 PDFAnonymous gguTkwGAAún no hay calificaciones
- Bioquimica ApuntesDocumento23 páginasBioquimica ApuntesDeejay JoseAún no hay calificaciones
- Semana 2 Acidos Nucleicos y El Flujo de La Informacion Genetica PDFDocumento86 páginasSemana 2 Acidos Nucleicos y El Flujo de La Informacion Genetica PDFRenato Gabriel Lopez AranaAún no hay calificaciones
- Biologia Selectividad Examen 3 Resuelto Castilla y Leon WWW - Siglo21x.blogspotDocumento7 páginasBiologia Selectividad Examen 3 Resuelto Castilla y Leon WWW - Siglo21x.blogspotmonografiasxAún no hay calificaciones
- 28 Ejercicios Resueltos de Energhia Calo PDFDocumento27 páginas28 Ejercicios Resueltos de Energhia Calo PDFjuanchiAún no hay calificaciones
- Biologia 2 Bachiller Sanchez Guillen PDFDocumento135 páginasBiologia 2 Bachiller Sanchez Guillen PDFSergio Salazar LatorreAún no hay calificaciones
- Tema 1. Formulación InorgánicaDocumento13 páginasTema 1. Formulación InorgánicaEstefania1987265Aún no hay calificaciones
- BIOLOGIA-SELECTIVIDAD-EXAMEN 9 RESUELTO-CATALUÑA-www - SIGLO21X.blogspotDocumento12 páginasBIOLOGIA-SELECTIVIDAD-EXAMEN 9 RESUELTO-CATALUÑA-www - SIGLO21X.blogspotmonografiasxAún no hay calificaciones
- Quiz Periodo 4to Noveno 1Documento3 páginasQuiz Periodo 4to Noveno 1Nikol LopezAún no hay calificaciones
- Cuadro Estructura ProteínasDocumento1 páginaCuadro Estructura ProteínasProf. Víctor M. VitoriaAún no hay calificaciones
- Esquema Clasificación y Función de Las ProteínasDocumento3 páginasEsquema Clasificación y Función de Las ProteínasProf. Víctor M. Vitoria100% (1)
- BYG 3 Esquema Del Tema 05 PDFDocumento7 páginasBYG 3 Esquema Del Tema 05 PDFrosa luzAún no hay calificaciones
- Selectividad Biologia PDFDocumento285 páginasSelectividad Biologia PDFFRANAún no hay calificaciones
- Biologia Selectividad Examen Resuelto Carabria 1cbs0lbiaec WWW - Siglo21x.blogspotDocumento7 páginasBiologia Selectividad Examen Resuelto Carabria 1cbs0lbiaec WWW - Siglo21x.blogspotmonografiasx100% (1)
- Formulacion Inorganica Diego Navarrete PDFDocumento29 páginasFormulacion Inorganica Diego Navarrete PDFJuan Pedreno BernalAún no hay calificaciones
- Tema 5. EnzimasDocumento10 páginasTema 5. EnzimasPablo VillaescusaAún no hay calificaciones
- Complejo Enzima-SustratoDocumento3 páginasComplejo Enzima-SustratoVERBEL PACHECO ANDRES FELIPEAún no hay calificaciones
- Biocatalizadores PDFDocumento93 páginasBiocatalizadores PDFJuan Camilo Gonzalez100% (1)
- Resumen EnzimasDocumento9 páginasResumen EnzimasOscarAún no hay calificaciones
- BiomoleculasDocumento11 páginasBiomoleculasYesenia Quiceno SernaAún no hay calificaciones
- Bio II Tema IIDocumento9 páginasBio II Tema IIbeabergamo87100% (1)
- Bio II Sec 04Documento3 páginasBio II Sec 04Luis Gerardo Pérez CastroAún no hay calificaciones
- Notas Bio II Seccion 02Documento6 páginasNotas Bio II Seccion 02beabergamo87Aún no hay calificaciones
- Notas Bio I Seccion 05 BionalisisDocumento2 páginasNotas Bio I Seccion 05 Bionalisisbeabergamo87Aún no hay calificaciones
- Conservación de Alimentos y Alimentos AlergenosDocumento66 páginasConservación de Alimentos y Alimentos AlergenosGina NuñezAún no hay calificaciones
- Tecnicas de Recoleccion de Insectos-With-Cover-Page-V2Documento25 páginasTecnicas de Recoleccion de Insectos-With-Cover-Page-V2Amare BeikerAún no hay calificaciones
- EstudiaDocumento16 páginasEstudiaLuciana Galindo0% (1)
- Resumen (Sistema Cardiovascular) .Documento6 páginasResumen (Sistema Cardiovascular) .angelicarosalescuy1Aún no hay calificaciones
- Mitosis y MeiosisDocumento8 páginasMitosis y MeiosisArnoldo MorónAún no hay calificaciones
- La Piel 3Documento15 páginasLa Piel 3Escuela Profesional DE Enfermeria UdchAún no hay calificaciones
- Alelos MúltiplesDocumento5 páginasAlelos MúltiplesMalicia VázquezAún no hay calificaciones
- 03 EcologíaDocumento4 páginas03 Ecologíajohn SnowAún no hay calificaciones
- Hidrobiología y Ecología de Los Microorganismos AcuáticosDocumento39 páginasHidrobiología y Ecología de Los Microorganismos AcuáticosJesus Ospino RiveraAún no hay calificaciones
- Reino FungiDocumento1 páginaReino FungiAlberto BrenesAún no hay calificaciones
- WDCDocumento555 páginasWDCRoberto MartinsAún no hay calificaciones
- Conceptualización de Las VariablesDocumento12 páginasConceptualización de Las VariablesZenaide MarcanoAún no hay calificaciones
- Album Gramineas y LeguminosasDocumento50 páginasAlbum Gramineas y LeguminosasAbdiel de Jesus Valencia HernandezAún no hay calificaciones
- Discurso Clínica Derechos de Los AnimalesDocumento4 páginasDiscurso Clínica Derechos de Los AnimalesAriadna G. Beroiz DíazAún no hay calificaciones
- Fluidos Corporales Investigacion CriminalDocumento16 páginasFluidos Corporales Investigacion CriminalFidel Aguirre CruzAún no hay calificaciones
- Pro. AgricolaDocumento41 páginasPro. Agricolacdsaantana561592Aún no hay calificaciones
- Jimenez Peyote Patrimonio Biocultural Mexico Sosteibilidad Tesis UG 2022Documento115 páginasJimenez Peyote Patrimonio Biocultural Mexico Sosteibilidad Tesis UG 2022interkevsAún no hay calificaciones
- Ensayo Sobre Tecnologia y CulturaDocumento4 páginasEnsayo Sobre Tecnologia y CulturaHenryAún no hay calificaciones
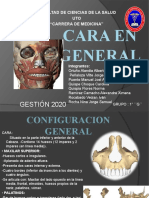
- Unidad V (Cara General) 1GDocumento22 páginasUnidad V (Cara General) 1GPeñaloza Vilte Jorge MartinAún no hay calificaciones
- 953am 5e8b508c91715BIODocumento8 páginas953am 5e8b508c91715BIORobert DietzAún no hay calificaciones
- Desarrollo y Crecimiento VegetalDocumento10 páginasDesarrollo y Crecimiento VegetalSilvana VillacortaAún no hay calificaciones
- GfytbfyDocumento5 páginasGfytbfyLeryeAún no hay calificaciones
- Manual de Extracción de Sangre en TortugasDocumento15 páginasManual de Extracción de Sangre en TortugasIsabel ContrerasAún no hay calificaciones
- Municipio ApizacoDocumento2 páginasMunicipio ApizacofamiliarmmAún no hay calificaciones