También podría gustarte
- Capas de La GeosferaDocumento3 páginasCapas de La GeosferaMiguel CandiaAún no hay calificaciones
- Taxonomia de Bloomklh.Documento2 páginasTaxonomia de Bloomklh.Miguel CandiaAún no hay calificaciones
- Fotos in Tes Is 2011Documento6 páginasFotos in Tes Is 2011Maria Jose Cuevas LopezAún no hay calificaciones
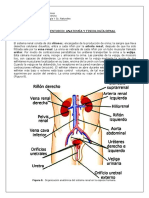
- 3° - Guía de Estudio - Anatomía y Fisiología RenalDocumento12 páginas3° - Guía de Estudio - Anatomía y Fisiología RenalMiguel CandiaAún no hay calificaciones
- U3 Experimentacion 2Documento4 páginasU3 Experimentacion 2Miguel CandiaAún no hay calificaciones
- 3º - Guía Estudio Higiene NerviosaDocumento10 páginas3º - Guía Estudio Higiene NerviosaMiguel CandiaAún no hay calificaciones
- Lista de MaterialesDocumento1 páginaLista de MaterialesMiguel CandiaAún no hay calificaciones
- Pruebas de Comprension Lectora 5 BasicoDocumento11 páginasPruebas de Comprension Lectora 5 BasicoAntonia AvalosAún no hay calificaciones
- Cuadro Con La Taxonomía de Bloom 03Documento5 páginasCuadro Con La Taxonomía de Bloom 03mauriciorvillalobos5993Aún no hay calificaciones
- Polímeros IIIDocumento2 páginasPolímeros IIIMiguel CandiaAún no hay calificaciones
- 3º - Guía Estudio Higiene NerviosaDocumento10 páginas3º - Guía Estudio Higiene NerviosaMiguel CandiaAún no hay calificaciones
- Guia Tipos Cel 3°electDocumento8 páginasGuia Tipos Cel 3°electMiguel CandiaAún no hay calificaciones
- Metodo Cientifico ExplicadoDocumento14 páginasMetodo Cientifico ExplicadoluisvaskoAún no hay calificaciones
- 3° - Ejercitación - HomeostasisDocumento3 páginas3° - Ejercitación - HomeostasisMiguel CandiaAún no hay calificaciones
- Potencial EsDocumento19 páginasPotencial EsMiguel CandiaAún no hay calificaciones
- Competencias Clave 1Documento13 páginasCompetencias Clave 1Miguel CandiaAún no hay calificaciones
- Regulacin de La Temperatura CorporalDocumento11 páginasRegulacin de La Temperatura CorporalMiguel CandiaAún no hay calificaciones
- Rúbrica para Evaluar El Reporte de InvestigaciónDocumento1 páginaRúbrica para Evaluar El Reporte de InvestigaciónMiguel CandiaAún no hay calificaciones
- Respuesta Guía MembranasDocumento5 páginasRespuesta Guía MembranasMiguel CandiaAún no hay calificaciones
- VirusDocumento20 páginasVirusMiguel CandiaAún no hay calificaciones
- HominizaciónDocumento1 páginaHominizaciónMiguel CandiaAún no hay calificaciones
- Plant AsDocumento17 páginasPlant AsMiguel CandiaAún no hay calificaciones
- El Sistema InmuneDocumento12 páginasEl Sistema InmuneAdriánSotoCuevasAún no hay calificaciones
- Guía 4º 2013Documento4 páginasGuía 4º 2013Miguel CandiaAún no hay calificaciones
- TALLER N°1 Quimica HidrocarburosDocumento3 páginasTALLER N°1 Quimica HidrocarburosMiguel CandiaAún no hay calificaciones
- Pauta de Evaluación de Exposición Oral Coeficiente 1Documento1 páginaPauta de Evaluación de Exposición Oral Coeficiente 1Miguel CandiaAún no hay calificaciones
- Evolucion Humana 3º Dif 2011Documento8 páginasEvolucion Humana 3º Dif 2011Miguel CandiaAún no hay calificaciones
- 3ro Guia N1 TermoquimicaDocumento2 páginas3ro Guia N1 TermoquimicaFaby AlbisturAún no hay calificaciones
- Enfermedades de La VisiónDocumento1 páginaEnfermedades de La VisiónMiguel CandiaAún no hay calificaciones
- Prueba Mañio InmuneDocumento5 páginasPrueba Mañio InmuneNatalia Carolina Osorio DuranAún no hay calificaciones
- Atención médica integral en el primer nivelDocumento114 páginasAtención médica integral en el primer nivelEfrain Antonio Jimenez MontenegroAún no hay calificaciones
- Ensayo Sobre El Trastorno BipolarDocumento13 páginasEnsayo Sobre El Trastorno BipolarElvis Manquez100% (1)
- Suturas6 3Documento8 páginasSuturas6 3andres_angel_50100% (1)
- Focusing DibujadoDocumento11 páginasFocusing DibujadoGonzalo Abarzúa100% (1)
- Etapas de La Reproducción: Unidad 3Documento13 páginasEtapas de La Reproducción: Unidad 3isabel carrilloAún no hay calificaciones
- Enlace QuímicoDocumento6 páginasEnlace QuímicoWilmer John Quincho CabreraAún no hay calificaciones
- Guia Informativa 8-1Documento3 páginasGuia Informativa 8-1CristhianAlexisFrancoEcheverriaAún no hay calificaciones
- S12 - Cavidad Nasal - Laringe - Traquea - PulmonDocumento39 páginasS12 - Cavidad Nasal - Laringe - Traquea - PulmonmelissaAún no hay calificaciones
- EcologíaDocumento2 páginasEcologíaEliza ZavaletaAún no hay calificaciones
- Paola Trabajo de La MicrobiataDocumento1 páginaPaola Trabajo de La MicrobiataSEGUNDO ELOY VALLEJOS BRAVOAún no hay calificaciones
- Lectura 2017-6 ZelekDocumento15 páginasLectura 2017-6 ZelekYolanda HernandezAún no hay calificaciones
- 1o 1T BIOLOGIA MAESTRO CUADERNILLO DE ACTIVIDADES 1TDocumento48 páginas1o 1T BIOLOGIA MAESTRO CUADERNILLO DE ACTIVIDADES 1TLuz Avila100% (2)
- Prevención MARSI y LESCAH: Enfoque inicial para lesiones cutáneasDocumento51 páginasPrevención MARSI y LESCAH: Enfoque inicial para lesiones cutáneasSantiago Estrada100% (1)
- CAPÍTULO I SistemasDocumento8 páginasCAPÍTULO I SistemasGustavo A. Puita ChoqueAún no hay calificaciones
- Prefijos GriegosDocumento5 páginasPrefijos GriegosMAUREENMONTANIAAún no hay calificaciones
- Resumen 3 Corte BiomolDocumento6 páginasResumen 3 Corte BiomolMarcela MuñozAún no hay calificaciones
- Lista de étimos y su significadoDocumento3 páginasLista de étimos y su significadoElizabeth InocAún no hay calificaciones
- Manual de Intervención en Crisis en Situaciones de Desastre PDFDocumento38 páginasManual de Intervención en Crisis en Situaciones de Desastre PDFDr. José Antonio Ayerdis ZamoraAún no hay calificaciones
- Test de Capurro: estimación de la edad gestacionalDocumento12 páginasTest de Capurro: estimación de la edad gestacionalGLORIA LOPEZAún no hay calificaciones
- BellisDocumento3 páginasBellisJANEAún no hay calificaciones
- Charla 5min JulioDocumento22 páginasCharla 5min JulioDiego Molina ChavezAún no hay calificaciones
- Concepto y Ambito de La PersonalidadDocumento41 páginasConcepto y Ambito de La PersonalidadMoises Nomeli Molina LopezAún no hay calificaciones
- Hawaian Olom I LomiDocumento5 páginasHawaian Olom I LomiLuis Romance Martinez100% (1)
- Capacitación exclusiva para profesionales de la estética: Adiposidad localizada, flaccidez cutánea y muscularDocumento29 páginasCapacitación exclusiva para profesionales de la estética: Adiposidad localizada, flaccidez cutánea y muscularVivian RigoAún no hay calificaciones
- Clase Oomycetes y Zygomycetes: Características y géneros de estos hongosDocumento17 páginasClase Oomycetes y Zygomycetes: Características y géneros de estos hongosDesarrollosgj CcaAún no hay calificaciones
- Resumen La Adolescencia Normal - Arminda AberasturyDocumento13 páginasResumen La Adolescencia Normal - Arminda AberasturyEugenio100% (2)
- ACTIVIDAD 1 Riesgos Psicosociales GlosarioDocumento14 páginasACTIVIDAD 1 Riesgos Psicosociales Glosariokatherine ruedaAún no hay calificaciones
- Exposición AgroclimatologíaDocumento15 páginasExposición AgroclimatologíaDaniel AlonsoAún no hay calificaciones
- Microscopia Electronica de TransmisionDocumento11 páginasMicroscopia Electronica de TransmisionMarcos LoredoAún no hay calificaciones
- La Conchuela o CoquinaDocumento11 páginasLa Conchuela o CoquinaDiego Manuel Torres SarzozoAún no hay calificaciones