También podría gustarte
- Citas (Andy Renato Flores Diaz)Documento10 páginasCitas (Andy Renato Flores Diaz)Antonio DiazAún no hay calificaciones
- Revision Bibliografica de MicropropagacionDocumento4 páginasRevision Bibliografica de MicropropagacionManuel SantanderAún no hay calificaciones
- Taller RegulacionDocumento4 páginasTaller RegulacionKevin España BurgosAún no hay calificaciones
- Ensayo de BiotecnologiaDocumento4 páginasEnsayo de BiotecnologiaStalin ProañoAún no hay calificaciones
- Ensayo de Gentica N02Documento10 páginasEnsayo de Gentica N02steven coralAún no hay calificaciones
- Micropropagación in Vitro de Nothofagus Alpina Utilizando FitohormonasDocumento10 páginasMicropropagación in Vitro de Nothofagus Alpina Utilizando FitohormonasStebanrojasAún no hay calificaciones
- Germoplasma 1161158274546Documento20 páginasGermoplasma 1161158274546JOSE LUIS ARTETA ALEGRÍAAún no hay calificaciones
- Interciencia 0378-1844: Issn: Interciencia@ivic - VeDocumento9 páginasInterciencia 0378-1844: Issn: Interciencia@ivic - Veanerak0412Aún no hay calificaciones
- La Crioterapia de ÁpicesDocumento13 páginasLa Crioterapia de ÁpicesNadir Eliecer Santos Chica100% (1)
- Melon - Trnsformacion GeneticaDocumento8 páginasMelon - Trnsformacion GeneticaAlejandro MilanoAún no hay calificaciones
- Proyecto Mariposas (Investigacion I)Documento4 páginasProyecto Mariposas (Investigacion I)daniel sierraAún no hay calificaciones
- BiotecnologiaDocumento90 páginasBiotecnologiaSayra Lizzett Vargas VillalobosAún no hay calificaciones
- Arroz DoradoDocumento17 páginasArroz DoradoHubert Sanchez ManayayAún no hay calificaciones
- Artículo Sobre La Conservación de Germoplasma de Especies Raras y Amenazadas (Revisión) Por J M Iriondo Alegría (2001)Documento20 páginasArtículo Sobre La Conservación de Germoplasma de Especies Raras y Amenazadas (Revisión) Por J M Iriondo Alegría (2001)ClaudioBertonattiAún no hay calificaciones
- Isaura MartinDocumento9 páginasIsaura MartinManuel HerreraAún no hay calificaciones
- Ensayo FitotecniaDocumento17 páginasEnsayo FitotecniaJoaquin CorreaAún no hay calificaciones
- Mejoramiento genético de plantas: Segunda EdiciónDe EverandMejoramiento genético de plantas: Segunda EdiciónCalificación: 3 de 5 estrellas3/5 (5)
- Efecto Del Miceliio Ganoderma LucidumDocumento7 páginasEfecto Del Miceliio Ganoderma LucidumJorge Salinas MoralesAún no hay calificaciones
- Manual para el cultivo de hortalizas. Familia SolanáceasDe EverandManual para el cultivo de hortalizas. Familia SolanáceasCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Plantas Transgenicas y Sus AplicacionesDocumento41 páginasPlantas Transgenicas y Sus AplicacionesIsaac ZapataAún no hay calificaciones
- Alimentos TransgenicosDocumento16 páginasAlimentos TransgenicosJessica AguirreAún no hay calificaciones
- Semillas Cultivos Alimenticios WilderDocumento6 páginasSemillas Cultivos Alimenticios WilderWilder Adolfo Cubas TantajulcaAún no hay calificaciones
- AgrobiodiversidadDocumento14 páginasAgrobiodiversidadGianFrancoG.GuibovichRamirezAún no hay calificaciones
- Proyecto BioestadísticaDocumento7 páginasProyecto BioestadísticaJesus Alexis Cervantes RamirezAún no hay calificaciones
- Cultivo in Vitro de P. Dubium y E. ContortisiliquumDocumento17 páginasCultivo in Vitro de P. Dubium y E. ContortisiliquumLarissa MeaurioAún no hay calificaciones
- Causas y Efectos Del Uso de Transgénicos en El Medio AmbienteDocumento12 páginasCausas y Efectos Del Uso de Transgénicos en El Medio AmbienteIvan BMAún no hay calificaciones
- Aplicaciones de La Reproducción Vegetativa (Johan Manuel Del Rosario S.) 26Documento4 páginasAplicaciones de La Reproducción Vegetativa (Johan Manuel Del Rosario S.) 26Johan SanchezAún no hay calificaciones
- LECTURA Microrganismos y BiodiversidadDocumento5 páginasLECTURA Microrganismos y BiodiversidadCam PosadasAún no hay calificaciones
- Reproducción Selectiva de Especies Forestales de Rapido CrecimientoDocumento68 páginasReproducción Selectiva de Especies Forestales de Rapido CrecimientoMarcelo Hernandez Nauto0% (1)
- Recursos NaturalesDocumento4 páginasRecursos NaturalesZaitu YuioAún no hay calificaciones
- Agro 2000 No. 87Documento51 páginasAgro 2000 No. 87Gustavo FloresAún no hay calificaciones
- Tesis GermoplasmaDocumento64 páginasTesis GermoplasmaCarlos RobertoAún no hay calificaciones
- MasificacionDocumento32 páginasMasificaciontefa sgAún no hay calificaciones
- La Deforestación y Sus CausasDocumento9 páginasLa Deforestación y Sus CausasAlejandro RamirezAún no hay calificaciones
- Proyecto Feria de Ciencias 2do Grado 2023Documento18 páginasProyecto Feria de Ciencias 2do Grado 2023Mariana Gomez100% (3)
- Propagacion de Plantas Nativos Por Sistema VegetativoDocumento28 páginasPropagacion de Plantas Nativos Por Sistema VegetativoCristian BarretoAún no hay calificaciones
- Articulo Enraizamiento Queñua 297-1502-1-PBDocumento8 páginasArticulo Enraizamiento Queñua 297-1502-1-PBEdwin BustamanteAún no hay calificaciones
- 84-Texto Del Artículo-181-1-10-20170222Documento12 páginas84-Texto Del Artículo-181-1-10-20170222Luis CruzAún no hay calificaciones
- Germoplasma VegetalDocumento18 páginasGermoplasma Vegetalprietalinda100% (1)
- Articulo Sobre El Arbol de La QuinaDocumento8 páginasArticulo Sobre El Arbol de La QuinaJoel Vilca PizarroAún no hay calificaciones
- Documento CompletoDocumento38 páginasDocumento CompletoMaria CandelaAún no hay calificaciones
- Agroforestería y BiodiversidadDocumento16 páginasAgroforestería y BiodiversidadJefferson Alexander Roriguez SoteloAún no hay calificaciones
- Clases GMF - 2013-IDocumento123 páginasClases GMF - 2013-IddddAún no hay calificaciones
- Biodiversidad y Aspectos Eticos y Morales de Los Cultivos TransgenicosDocumento26 páginasBiodiversidad y Aspectos Eticos y Morales de Los Cultivos TransgenicosNataly Valverde RicraAún no hay calificaciones
- La BiodiversidadDocumento13 páginasLa BiodiversidadJorge CoronaAún no hay calificaciones
- Produccion - de - Semillas MoringaDocumento9 páginasProduccion - de - Semillas MoringaCesar MurgaAún no hay calificaciones
- Valoración Económica de La PolinizaciónDocumento7 páginasValoración Económica de La PolinizaciónMICROBIOPERU100% (1)
- Los Organismos Genéticamente Modificados y El Medio AmbienteDocumento9 páginasLos Organismos Genéticamente Modificados y El Medio AmbienteAngie Catalina Guerrero LopezAún no hay calificaciones
- Mejoramiento Genetico y Biotecnologico de Plantas-LibreDocumento286 páginasMejoramiento Genetico y Biotecnologico de Plantas-LibreJennifer Baker100% (1)
- Documento CompletoDocumento6 páginasDocumento CompletoEster EncizoAún no hay calificaciones
- Proyecto BioestadísticaDocumento8 páginasProyecto BioestadísticaJesus Alexis Cervantes RamirezAún no hay calificaciones
- Epi - Coronel - RamosDocumento17 páginasEpi - Coronel - RamosAna AguilarAún no hay calificaciones
- Evaluación de Variedades Nativas y Mejoradas Bajo El Sistema MilpaDocumento6 páginasEvaluación de Variedades Nativas y Mejoradas Bajo El Sistema MilpaanyAún no hay calificaciones
- Biod Sarandon 2020Documento27 páginasBiod Sarandon 2020Dany BalanzarAún no hay calificaciones
- Bancos Mixtos Forrajeros Como Estrategia para El Rescate de Semillas e Impulso de La EndogenológicaDocumento10 páginasBancos Mixtos Forrajeros Como Estrategia para El Rescate de Semillas e Impulso de La Endogenológicabernalastorgam1Aún no hay calificaciones
- Bfpa U4 A2 VabcDocumento5 páginasBfpa U4 A2 VabcVane SchatzAún no hay calificaciones
- REQUERIMIENTOSDocumento12 páginasREQUERIMIENTOSPaulo Rodríguez RomeroAún no hay calificaciones
- Tarea-2 - Importancia-Economica-y-Categorizacion-TaxonomicaDocumento14 páginasTarea-2 - Importancia-Economica-y-Categorizacion-Taxonomicayesica lorena villa blandonAún no hay calificaciones
- Utilizacionbiorresiduos PDFDocumento12 páginasUtilizacionbiorresiduos PDFvictor moralesAún no hay calificaciones
- Encapsulación de probióticos: Alternativa tecnológica para mejorar el cultivo de tilapiaDe EverandEncapsulación de probióticos: Alternativa tecnológica para mejorar el cultivo de tilapiaAún no hay calificaciones
- Valores de Referencia para Las Pruebas de Coagulacion en MexicoDocumento7 páginasValores de Referencia para Las Pruebas de Coagulacion en MexicoAlfredo Capo PfmAún no hay calificaciones
- 2005 2006 Edu 08 TemaDocumento9 páginas2005 2006 Edu 08 TemaDinora MillaAún no hay calificaciones
- Factores de Virulencia Bacterianos y PatogenicidadDocumento42 páginasFactores de Virulencia Bacterianos y PatogenicidadAlfredo Capo PfmAún no hay calificaciones
- Estructura y Función Celular de Los Microorganismos.Documento15 páginasEstructura y Función Celular de Los Microorganismos.Alfredo Capo PfmAún no hay calificaciones
- ANTICOAGULANTESDocumento4 páginasANTICOAGULANTESAlfredo Capo Pfm100% (1)
- Autoclaves de LaboratorioDocumento22 páginasAutoclaves de LaboratorioAlfredo Capo PfmAún no hay calificaciones
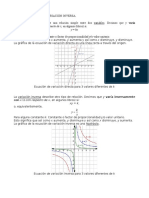
- Variación Directa y Variación InversaDocumento3 páginasVariación Directa y Variación InversaChilo_57100% (1)
- Historia de La Provincia de NeuquenDocumento129 páginasHistoria de La Provincia de NeuquenClaudia Ormaechea RussoAún no hay calificaciones
- KatididoDocumento3 páginasKatididoClaudio Alejandro Chavez MoscosoAún no hay calificaciones
- Entomologia ChaparroDocumento9 páginasEntomologia ChaparroOswaldo ARAún no hay calificaciones
- Cuadernillo Nº7 Abonos OrgánicosDocumento34 páginasCuadernillo Nº7 Abonos OrgánicosCorporación Ecológica y Cultural Penca de Sábila100% (1)
- Proyecto de NaranjaDocumento47 páginasProyecto de NaranjaRenzo CastillaAún no hay calificaciones
- 111 Platos Populares Del EcuadorDocumento21 páginas111 Platos Populares Del EcuadorDavid Quinteros C.Aún no hay calificaciones
- Elaboracion de Chrorizo TradicionalDocumento58 páginasElaboracion de Chrorizo TradicionalEdwin J. MartinezAún no hay calificaciones
- Estudio ComparativoDocumento3 páginasEstudio ComparativoJhorman Alexis Quintero100% (1)
- CAPÍTULO 3 Establecimiento Del Cultivo de ArandanoDocumento2 páginasCAPÍTULO 3 Establecimiento Del Cultivo de ArandanoKarin Santana GonzalesAún no hay calificaciones
- Cuadro ComparativoDocumento17 páginasCuadro ComparativoKarla ZelayaAún no hay calificaciones
- PolloDocumento22 páginasPolloPaolitha TorresAún no hay calificaciones
- Recursos Forestales Parte 1Documento32 páginasRecursos Forestales Parte 1Diligenciamientos Matilde NietoAún no hay calificaciones
- Danzas de La LibertadDocumento4 páginasDanzas de La LibertadJulio Hamilton50% (2)
- REGION de CoquimboDocumento6 páginasREGION de CoquimboantaAún no hay calificaciones
- Formulario Numeros IndicesDocumento4 páginasFormulario Numeros IndicesAndre Edy Ramos PeñaAún no hay calificaciones
- Catalogo de Productos Forestales PDFDocumento34 páginasCatalogo de Productos Forestales PDFZareth GarfiazzAún no hay calificaciones
- Cuero Piel TilapiaDocumento72 páginasCuero Piel TilapiaCamelo AnggyeAún no hay calificaciones
- Triptico El VirreynatoDocumento2 páginasTriptico El VirreynatoLuis TeránAún no hay calificaciones
- Mineria y Las BambasDocumento78 páginasMineria y Las BambasvalentincondoriAún no hay calificaciones
- Planteamiento Del Problema:: ProgramacionDocumento8 páginasPlanteamiento Del Problema:: ProgramacionDafne LopézAún no hay calificaciones
- PDN (Cuyes) - Apea AijaDocumento76 páginasPDN (Cuyes) - Apea AijaRoger Maradona Morales LazaroAún no hay calificaciones
- Pd-Introduccion y Antecedentes - El Santuario - AntioquiaDocumento31 páginasPd-Introduccion y Antecedentes - El Santuario - AntioquiaSU CASA ES UNA ESCUELA DE ARTEAún no hay calificaciones
- Plan Negocios Compost Final CUYAZA PDFDocumento79 páginasPlan Negocios Compost Final CUYAZA PDFWalter Silvera RiveraAún no hay calificaciones
- La Rosa Más BellaDocumento7 páginasLa Rosa Más BellaLucia CoronelAún no hay calificaciones
- BPA - Metodología para Su ImplementaciónDocumento128 páginasBPA - Metodología para Su ImplementaciónAlberto Miguel Pastor Patiño100% (1)
- Alm. Sum. - DRDocumento7 páginasAlm. Sum. - DRAnonymous imscLyAún no hay calificaciones
- Engeo 247 ZCDocumento8 páginasEngeo 247 ZCBraulio Jimenez BravoAún no hay calificaciones
- Proyecto de Producción de FloresDocumento19 páginasProyecto de Producción de FloresLeo AquinoAún no hay calificaciones