
Dolor
Dolor
También podría gustarte
- Anatomia y Fisiología del Sistema Nervioso IIDe EverandAnatomia y Fisiología del Sistema Nervioso IICalificación: 4 de 5 estrellas4/5 (28)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- PletoDocumento112 páginasPletoemmamarty97% (39)
- Fisiologia Del DolorDocumento38 páginasFisiologia Del DolorFrancisco Javier Palma PAún no hay calificaciones
- Fisiopatologia Del DolorDocumento34 páginasFisiopatologia Del DolorAnto Flores100% (4)
- Neurofisiologia Del Dolor 2014Documento31 páginasNeurofisiologia Del Dolor 2014Guido Orlando100% (1)
- Los Cuatro Procesos Fisiológicos en El DolorDocumento6 páginasLos Cuatro Procesos Fisiológicos en El Dolorsusana_sanzana100% (1)
- Anatomía y Fisiología del Sistema NerviosoDe EverandAnatomía y Fisiología del Sistema NerviosoCalificación: 4 de 5 estrellas4/5 (19)
- Protocolo de Duran y Houser122Documento12 páginasProtocolo de Duran y Houser122Annie100% (2)
- Neuroatomía Del DolorDocumento6 páginasNeuroatomía Del DolorUkko RannikkoAún no hay calificaciones
- Neuropsicología Del DolorDocumento7 páginasNeuropsicología Del DolorMichael Steven Buesaquillo PAún no hay calificaciones
- Fisiologia Del DolorDocumento7 páginasFisiologia Del DolorCris Tian Ignacio AstudilloAún no hay calificaciones
- Fisiologia Del DolorDocumento14 páginasFisiologia Del DolorGeremias Ojeda Huaman100% (1)
- Fisiopatologia Del DolorDocumento15 páginasFisiopatologia Del DolorOmar Bazaldua100% (1)
- Guia Neurofisiologia Del Dolor Daniel PatinoDocumento6 páginasGuia Neurofisiologia Del Dolor Daniel PatinoAndrés ContrerasAún no hay calificaciones
- FisiopatodolorDocumento14 páginasFisiopatodolorJuanJoséCastilloHinojosaAún no hay calificaciones
- S3 Neurofisiologia I ParDocumento9 páginasS3 Neurofisiologia I ParZullyAún no hay calificaciones
- CW Es Ec Dolor NeuroDocumento18 páginasCW Es Ec Dolor NeuroHelenaAún no hay calificaciones
- J Mpaic 2013 08 004 en EsDocumento4 páginasJ Mpaic 2013 08 004 en EsholguinlAún no hay calificaciones
- Fisiopatología Del DolorDocumento42 páginasFisiopatología Del DolorsabeduAún no hay calificaciones
- T-1.1 Generalidades, Dolor 2020-2Documento75 páginasT-1.1 Generalidades, Dolor 2020-2Erik J. Roshan FloresAún no hay calificaciones
- Dolor Lmixto Cambiando ParadigmasDocumento9 páginasDolor Lmixto Cambiando ParadigmasSindy AguilarAún no hay calificaciones
- Vias Centrales Del DolorDocumento5 páginasVias Centrales Del DolorEdher Lima GarciaAún no hay calificaciones
- Anatomia Basica y Fisiologia Del Dolor CaminosDocumento10 páginasAnatomia Basica y Fisiologia Del Dolor CaminosrosangelaAún no hay calificaciones
- T-1.1 Generalidades, DolorDocumento113 páginasT-1.1 Generalidades, DolorBryan Ripa ContrerasAún no hay calificaciones
- Monografia de Morfofisiologia (Grupo5)Documento20 páginasMonografia de Morfofisiologia (Grupo5)Jesús OlivaresAún no hay calificaciones
- Fisiopatologia Del Dolor PDFDocumento12 páginasFisiopatologia Del Dolor PDFDiego Gaston EnrichAún no hay calificaciones
- FISIOLOGÍA DEL DOLOR PARA EL CURSO 2021.en - EsDocumento5 páginasFISIOLOGÍA DEL DOLOR PARA EL CURSO 2021.en - EsMauricio MarínAún no hay calificaciones
- Dolor AgudoDocumento18 páginasDolor AgudoAide Gonzalez SandovalAún no hay calificaciones
- Tronco Encefalico InformeDocumento56 páginasTronco Encefalico InformeJackelyn Rimarachin Casique50% (2)
- Seminario 4 Grupo 1 Dolor Inflamacion No OpiaceosDocumento22 páginasSeminario 4 Grupo 1 Dolor Inflamacion No OpiaceosSabrina Reyna FarjeAún no hay calificaciones
- Vías Del Dolor AgudoDocumento4 páginasVías Del Dolor AgudoMARIA MAGDALENA ROACHO PICONAún no hay calificaciones
- Psicosomatica Del Dolor FinalDocumento10 páginasPsicosomatica Del Dolor FinalJoshep SangarAún no hay calificaciones
- Neurobiologia Del Dolor PDFDocumento16 páginasNeurobiologia Del Dolor PDFmanuel_matasAún no hay calificaciones
- DolorDocumento52 páginasDolorPercy Rodríguez Bravo100% (3)
- Dolor 3Documento95 páginasDolor 3Fabiola RodriguezAún no hay calificaciones
- Fisiopatologia Del DolorDocumento7 páginasFisiopatologia Del DolorRami CoRioAún no hay calificaciones
- Terminaciones NerviosasDocumento11 páginasTerminaciones Nerviosasdania camachoAún no hay calificaciones
- Neurofisiologia Del DolorDocumento23 páginasNeurofisiologia Del DolorAna GiraldoAún no hay calificaciones
- Fisiopatologia Del Dolor Porth 84465 Downloable 3158457Documento8 páginasFisiopatologia Del Dolor Porth 84465 Downloable 3158457Marcelo AlvarezAún no hay calificaciones
- Péptidos Opioides Endógenos, Dolor y Adicción: Revisión BibliográficaDocumento7 páginasPéptidos Opioides Endógenos, Dolor y Adicción: Revisión BibliográficaMiguel De Jesus Gallegos PeraltaAún no hay calificaciones
- Copia de Dolor ArgenteDocumento3 páginasCopia de Dolor ArgenteNATALIA FERNANDA SÁEZ ANTILEFAún no hay calificaciones
- Farmacología Del Dolor y La AnestesiaDocumento4 páginasFarmacología Del Dolor y La AnestesiaDaylis YepezAún no hay calificaciones
- Vías y Mecanismos de Transmisión Del Dolor NociceptoresDocumento2 páginasVías y Mecanismos de Transmisión Del Dolor NociceptoresCristopher Castro RdAún no hay calificaciones
- Opioid EsDocumento5 páginasOpioid EsSHARON LENNY MAMANI TONCONIAún no hay calificaciones
- FISIOPATOLOGIA DEL DOLOR - OkDocumento62 páginasFISIOPATOLOGIA DEL DOLOR - OkRainier PCAún no hay calificaciones
- S02.s2 - Material de ClaseDocumento22 páginasS02.s2 - Material de Claseingrid shirley polo ponteAún no hay calificaciones
- Notas de NocicepciónDocumento9 páginasNotas de NocicepciónCarolquiAún no hay calificaciones
- DolorDocumento18 páginasDolorVivi CardonaAún no hay calificaciones
- Neurofisiologia Del DolorDocumento21 páginasNeurofisiologia Del DolorLety Catari100% (1)
- Argente DolorDocumento29 páginasArgente DolorDAMARIS CRISTELL AGUILAR SANCHEZAún no hay calificaciones
- 2.1.4 Fisiología Del DolorDocumento3 páginas2.1.4 Fisiología Del DolorJosé ZamoraAún no hay calificaciones
- Dolorr FisiopatologiaDocumento16 páginasDolorr FisiopatologiaJAIRO GALINDOAún no hay calificaciones
- BASES FISIOPATOLÓGICAS DEL DOLOR Acta Médica Peruana, Mayo-Agosto, Año/vol. 24, Número 002Documento5 páginasBASES FISIOPATOLÓGICAS DEL DOLOR Acta Médica Peruana, Mayo-Agosto, Año/vol. 24, Número 002Paloma MendozaAún no hay calificaciones
- Practica Nro 3 Sistema Nervioso (24970)Documento4 páginasPractica Nro 3 Sistema Nervioso (24970)Gabriela GutierrezAún no hay calificaciones
- Dolor PDFDocumento39 páginasDolor PDFSaandy GC100% (1)
- Anatomía y Fisiología de La NocicepciónDocumento32 páginasAnatomía y Fisiología de La NocicepciónAndy OrtizAún no hay calificaciones
- Wa0104.Documento19 páginasWa0104.grupodeamistad20Aún no hay calificaciones
- Anatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2De EverandAnatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Aún no hay calificaciones
- El cannabis en patologías del sistema nervioso centralDe EverandEl cannabis en patologías del sistema nervioso centralAún no hay calificaciones
- Uso del cannabis en el manejo del dolorDe EverandUso del cannabis en el manejo del dolorCalificación: 3 de 5 estrellas3/5 (1)
- Estiramientos Mio-Tendino - Aponeuroticos 2 PDFDocumento12 páginasEstiramientos Mio-Tendino - Aponeuroticos 2 PDFAnnieAún no hay calificaciones
- Conformación Del Comité de BioéticaDocumento1 páginaConformación Del Comité de BioéticaAnnieAún no hay calificaciones
- NEUROFACILITACIÓNDocumento6 páginasNEUROFACILITACIÓNAnnieAún no hay calificaciones
- Extracto QUIROZ Humero Radio CubitoDocumento12 páginasExtracto QUIROZ Humero Radio CubitoAnnieAún no hay calificaciones








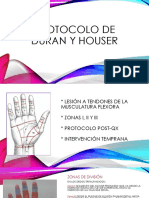
























































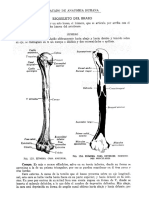
También podría gustarte
- Anatomia y Fisiología del Sistema Nervioso IIDe EverandAnatomia y Fisiología del Sistema Nervioso IICalificación: 4 de 5 estrellas4/5 (28)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)De EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Calificación: 5 de 5 estrellas5/5 (9)
- PletoDocumento112 páginasPletoemmamarty97% (39)
- Fisiologia Del DolorDocumento38 páginasFisiologia Del DolorFrancisco Javier Palma PAún no hay calificaciones
- Fisiopatologia Del DolorDocumento34 páginasFisiopatologia Del DolorAnto Flores100% (4)
- Neurofisiologia Del Dolor 2014Documento31 páginasNeurofisiologia Del Dolor 2014Guido Orlando100% (1)
- Los Cuatro Procesos Fisiológicos en El DolorDocumento6 páginasLos Cuatro Procesos Fisiológicos en El Dolorsusana_sanzana100% (1)
- Anatomía y Fisiología del Sistema NerviosoDe EverandAnatomía y Fisiología del Sistema NerviosoCalificación: 4 de 5 estrellas4/5 (19)
- Protocolo de Duran y Houser122Documento12 páginasProtocolo de Duran y Houser122Annie100% (2)
- Neuroatomía Del DolorDocumento6 páginasNeuroatomía Del DolorUkko RannikkoAún no hay calificaciones
- Neuropsicología Del DolorDocumento7 páginasNeuropsicología Del DolorMichael Steven Buesaquillo PAún no hay calificaciones
- Fisiologia Del DolorDocumento7 páginasFisiologia Del DolorCris Tian Ignacio AstudilloAún no hay calificaciones
- Fisiologia Del DolorDocumento14 páginasFisiologia Del DolorGeremias Ojeda Huaman100% (1)
- Fisiopatologia Del DolorDocumento15 páginasFisiopatologia Del DolorOmar Bazaldua100% (1)
- Guia Neurofisiologia Del Dolor Daniel PatinoDocumento6 páginasGuia Neurofisiologia Del Dolor Daniel PatinoAndrés ContrerasAún no hay calificaciones
- FisiopatodolorDocumento14 páginasFisiopatodolorJuanJoséCastilloHinojosaAún no hay calificaciones
- S3 Neurofisiologia I ParDocumento9 páginasS3 Neurofisiologia I ParZullyAún no hay calificaciones
- CW Es Ec Dolor NeuroDocumento18 páginasCW Es Ec Dolor NeuroHelenaAún no hay calificaciones
- J Mpaic 2013 08 004 en EsDocumento4 páginasJ Mpaic 2013 08 004 en EsholguinlAún no hay calificaciones
- Fisiopatología Del DolorDocumento42 páginasFisiopatología Del DolorsabeduAún no hay calificaciones
- T-1.1 Generalidades, Dolor 2020-2Documento75 páginasT-1.1 Generalidades, Dolor 2020-2Erik J. Roshan FloresAún no hay calificaciones
- Dolor Lmixto Cambiando ParadigmasDocumento9 páginasDolor Lmixto Cambiando ParadigmasSindy AguilarAún no hay calificaciones
- Vias Centrales Del DolorDocumento5 páginasVias Centrales Del DolorEdher Lima GarciaAún no hay calificaciones
- Anatomia Basica y Fisiologia Del Dolor CaminosDocumento10 páginasAnatomia Basica y Fisiologia Del Dolor CaminosrosangelaAún no hay calificaciones
- T-1.1 Generalidades, DolorDocumento113 páginasT-1.1 Generalidades, DolorBryan Ripa ContrerasAún no hay calificaciones
- Monografia de Morfofisiologia (Grupo5)Documento20 páginasMonografia de Morfofisiologia (Grupo5)Jesús OlivaresAún no hay calificaciones
- Fisiopatologia Del Dolor PDFDocumento12 páginasFisiopatologia Del Dolor PDFDiego Gaston EnrichAún no hay calificaciones
- FISIOLOGÍA DEL DOLOR PARA EL CURSO 2021.en - EsDocumento5 páginasFISIOLOGÍA DEL DOLOR PARA EL CURSO 2021.en - EsMauricio MarínAún no hay calificaciones
- Dolor AgudoDocumento18 páginasDolor AgudoAide Gonzalez SandovalAún no hay calificaciones
- Tronco Encefalico InformeDocumento56 páginasTronco Encefalico InformeJackelyn Rimarachin Casique50% (2)
- Seminario 4 Grupo 1 Dolor Inflamacion No OpiaceosDocumento22 páginasSeminario 4 Grupo 1 Dolor Inflamacion No OpiaceosSabrina Reyna FarjeAún no hay calificaciones
- Vías Del Dolor AgudoDocumento4 páginasVías Del Dolor AgudoMARIA MAGDALENA ROACHO PICONAún no hay calificaciones
- Psicosomatica Del Dolor FinalDocumento10 páginasPsicosomatica Del Dolor FinalJoshep SangarAún no hay calificaciones
- Neurobiologia Del Dolor PDFDocumento16 páginasNeurobiologia Del Dolor PDFmanuel_matasAún no hay calificaciones
- DolorDocumento52 páginasDolorPercy Rodríguez Bravo100% (3)
- Dolor 3Documento95 páginasDolor 3Fabiola RodriguezAún no hay calificaciones
- Fisiopatologia Del DolorDocumento7 páginasFisiopatologia Del DolorRami CoRioAún no hay calificaciones
- Terminaciones NerviosasDocumento11 páginasTerminaciones Nerviosasdania camachoAún no hay calificaciones
- Neurofisiologia Del DolorDocumento23 páginasNeurofisiologia Del DolorAna GiraldoAún no hay calificaciones
- Fisiopatologia Del Dolor Porth 84465 Downloable 3158457Documento8 páginasFisiopatologia Del Dolor Porth 84465 Downloable 3158457Marcelo AlvarezAún no hay calificaciones
- Péptidos Opioides Endógenos, Dolor y Adicción: Revisión BibliográficaDocumento7 páginasPéptidos Opioides Endógenos, Dolor y Adicción: Revisión BibliográficaMiguel De Jesus Gallegos PeraltaAún no hay calificaciones
- Copia de Dolor ArgenteDocumento3 páginasCopia de Dolor ArgenteNATALIA FERNANDA SÁEZ ANTILEFAún no hay calificaciones
- Farmacología Del Dolor y La AnestesiaDocumento4 páginasFarmacología Del Dolor y La AnestesiaDaylis YepezAún no hay calificaciones
- Vías y Mecanismos de Transmisión Del Dolor NociceptoresDocumento2 páginasVías y Mecanismos de Transmisión Del Dolor NociceptoresCristopher Castro RdAún no hay calificaciones
- Opioid EsDocumento5 páginasOpioid EsSHARON LENNY MAMANI TONCONIAún no hay calificaciones
- FISIOPATOLOGIA DEL DOLOR - OkDocumento62 páginasFISIOPATOLOGIA DEL DOLOR - OkRainier PCAún no hay calificaciones
- S02.s2 - Material de ClaseDocumento22 páginasS02.s2 - Material de Claseingrid shirley polo ponteAún no hay calificaciones
- Notas de NocicepciónDocumento9 páginasNotas de NocicepciónCarolquiAún no hay calificaciones
- DolorDocumento18 páginasDolorVivi CardonaAún no hay calificaciones
- Neurofisiologia Del DolorDocumento21 páginasNeurofisiologia Del DolorLety Catari100% (1)
- Argente DolorDocumento29 páginasArgente DolorDAMARIS CRISTELL AGUILAR SANCHEZAún no hay calificaciones
- 2.1.4 Fisiología Del DolorDocumento3 páginas2.1.4 Fisiología Del DolorJosé ZamoraAún no hay calificaciones
- Dolorr FisiopatologiaDocumento16 páginasDolorr FisiopatologiaJAIRO GALINDOAún no hay calificaciones
- BASES FISIOPATOLÓGICAS DEL DOLOR Acta Médica Peruana, Mayo-Agosto, Año/vol. 24, Número 002Documento5 páginasBASES FISIOPATOLÓGICAS DEL DOLOR Acta Médica Peruana, Mayo-Agosto, Año/vol. 24, Número 002Paloma MendozaAún no hay calificaciones
- Practica Nro 3 Sistema Nervioso (24970)Documento4 páginasPractica Nro 3 Sistema Nervioso (24970)Gabriela GutierrezAún no hay calificaciones
- Dolor PDFDocumento39 páginasDolor PDFSaandy GC100% (1)
- Anatomía y Fisiología de La NocicepciónDocumento32 páginasAnatomía y Fisiología de La NocicepciónAndy OrtizAún no hay calificaciones
- Wa0104.Documento19 páginasWa0104.grupodeamistad20Aún no hay calificaciones
- Anatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2De EverandAnatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Aún no hay calificaciones
- El cannabis en patologías del sistema nervioso centralDe EverandEl cannabis en patologías del sistema nervioso centralAún no hay calificaciones
- Uso del cannabis en el manejo del dolorDe EverandUso del cannabis en el manejo del dolorCalificación: 3 de 5 estrellas3/5 (1)
- Estiramientos Mio-Tendino - Aponeuroticos 2 PDFDocumento12 páginasEstiramientos Mio-Tendino - Aponeuroticos 2 PDFAnnieAún no hay calificaciones
- Conformación Del Comité de BioéticaDocumento1 páginaConformación Del Comité de BioéticaAnnieAún no hay calificaciones
- NEUROFACILITACIÓNDocumento6 páginasNEUROFACILITACIÓNAnnieAún no hay calificaciones
- Extracto QUIROZ Humero Radio CubitoDocumento12 páginasExtracto QUIROZ Humero Radio CubitoAnnieAún no hay calificaciones