También podría gustarte
- Josselyn Nora Nidia Esquite Montoya: Guatemala, Octubre de 2,016Documento50 páginasJosselyn Nora Nidia Esquite Montoya: Guatemala, Octubre de 2,016Luisa RamirezAún no hay calificaciones
- Romero Alcaraz IsmaelDocumento54 páginasRomero Alcaraz IsmaelLii GutierrezAún no hay calificaciones
- Universidad Mayor de San Andrés Facultad de Agronomía Carrera de Ingeniería AgronómicaDocumento101 páginasUniversidad Mayor de San Andrés Facultad de Agronomía Carrera de Ingeniería AgronómicaGustavo Cano GallegosAún no hay calificaciones
- 1 Pargo ColoradoDocumento84 páginas1 Pargo ColoradoErickRamirezFiestasAún no hay calificaciones
- Madera de MadidiDocumento141 páginasMadera de Madidi_i_o_Aún no hay calificaciones
- Arista Bustamante, Jessy PatriciaDocumento110 páginasArista Bustamante, Jessy PatriciaAmerica LeireAún no hay calificaciones
- Informe Zoocriaderos de Avestruces y MadDocumento19 páginasInforme Zoocriaderos de Avestruces y Madyovanny antonAún no hay calificaciones
- t16494 Solis Rosales, Adalberto TesisDocumento81 páginast16494 Solis Rosales, Adalberto TesisElias GutierrezAún no hay calificaciones
- Art-Bacterias Patogenas Agua PDFDocumento66 páginasArt-Bacterias Patogenas Agua PDFFer EspinozaAún no hay calificaciones
- Atlas Helmintos PDFDocumento182 páginasAtlas Helmintos PDFObarduckAún no hay calificaciones
- Atlas de HelmintosDocumento153 páginasAtlas de HelmintosGutierrez AleAún no hay calificaciones
- COL Series 3 PDFDocumento314 páginasCOL Series 3 PDFVanessa AlejandraAún no hay calificaciones
- Tesis Reproduccion Asexualde Espina de MarDocumento76 páginasTesis Reproduccion Asexualde Espina de Marangel100% (1)
- T033 45782767 TDocumento114 páginasT033 45782767 TBrayan BONILLA YAUYOAún no hay calificaciones
- Universidad Mayor de San AndresDocumento131 páginasUniversidad Mayor de San AndresJUAN CARLOS VARGAS APAZAAún no hay calificaciones
- Taxonomía y Filogenia de Trichocereus PDFDocumento252 páginasTaxonomía y Filogenia de Trichocereus PDFFred M Thompson0% (1)
- Manglares MonografiaDocumento107 páginasManglares MonografiaDanna Maricarmen Juarez GarciaAún no hay calificaciones
- Universidad Autónoma de Nuevo León: Facultad de Agronomía Facultad de Medicina Veterinaria Y ZootecniaDocumento266 páginasUniversidad Autónoma de Nuevo León: Facultad de Agronomía Facultad de Medicina Veterinaria Y Zootecniamauricior_789Aún no hay calificaciones
- AGamccbe033 PDFDocumento77 páginasAGamccbe033 PDFMauro GarcíaAún no hay calificaciones
- DESCRIPCION HISTOLOGICA DEL SISTEMA REPRODUCTOR DE LA HEMBRA DE CHUCHA COMUN (Djdelphis Maraupialis) PDF PDFDocumento51 páginasDESCRIPCION HISTOLOGICA DEL SISTEMA REPRODUCTOR DE LA HEMBRA DE CHUCHA COMUN (Djdelphis Maraupialis) PDF PDFKevindaniel CoVaAún no hay calificaciones
- Area Agropecuaria Y de Recursos Naturales RenovablesDocumento74 páginasArea Agropecuaria Y de Recursos Naturales Renovables6513joviAún no hay calificaciones
- BIapabrjDocumento79 páginasBIapabrjESPINOZA DIAZ GIANMARCOSAún no hay calificaciones
- Diseño Randomizado PDFDocumento56 páginasDiseño Randomizado PDFGladys Tanta ChugnasAún no hay calificaciones
- Lista Roja de Los Anfibios y Reptiles Del UruguayDocumento66 páginasLista Roja de Los Anfibios y Reptiles Del UruguayKarumbéAún no hay calificaciones
- Tesis Trips GuatemalaDocumento57 páginasTesis Trips GuatemalaVilma PorresAún no hay calificaciones
- Atlas y Libro Rojo de Los Mamiferos Terrestres de Espana PDFDocumento590 páginasAtlas y Libro Rojo de Los Mamiferos Terrestres de Espana PDFIgnacio Jisus Mendoza SchifferliAún no hay calificaciones
- Pulgas de Perros RuralesDocumento77 páginasPulgas de Perros RuralesIsa Yael Palacios Quevedo 6836Aún no hay calificaciones
- Ibader Pecuaria MY-683Documento64 páginasIbader Pecuaria MY-683DianaAún no hay calificaciones
- 10 VariedadesDocumento113 páginas10 VariedadesFederico Condori MontañoAún no hay calificaciones
- Tesis MV Maria Alejandra Gonzalez PDocumento43 páginasTesis MV Maria Alejandra Gonzalez Praul vera cuellarAún no hay calificaciones
- Diversidada de Mariposas El Salvador, Parque Walter Thilo DeinningerDocumento112 páginasDiversidada de Mariposas El Salvador, Parque Walter Thilo DeinningerRuben Sorto100% (3)
- PesqueríaDocumento19 páginasPesqueríaCintia ChabaAún no hay calificaciones
- Universidad Mayor de San Andrés Facultad de Agronomía Carrera de Ingeniería AgronómicaDocumento119 páginasUniversidad Mayor de San Andrés Facultad de Agronomía Carrera de Ingeniería AgronómicaJostin JoutexAún no hay calificaciones
- Cultivo de Jengibre 1Documento82 páginasCultivo de Jengibre 1Cyspagro de la CuencaAún no hay calificaciones
- Universidad Mayor de San Andrés Facultad de Agronomía Carrera de Ingeniería AgronómicaDocumento100 páginasUniversidad Mayor de San Andrés Facultad de Agronomía Carrera de Ingeniería Agronómicapablonahumquispe2024Aún no hay calificaciones
- EFECTOS DE LAS DENSIDADES POBLACIONALES Y LOS NIVELES DE FERTILIZACIÓN SOBRE EL RENDIMIENTOS DEL CULTIVO MANÍ (Arachis Hypogaea L)Documento65 páginasEFECTOS DE LAS DENSIDADES POBLACIONALES Y LOS NIVELES DE FERTILIZACIÓN SOBRE EL RENDIMIENTOS DEL CULTIVO MANÍ (Arachis Hypogaea L)Ender HerreraAún no hay calificaciones
- Berrendo DinamicaDocumento64 páginasBerrendo DinamicaarbolAún no hay calificaciones
- PelayoDocumento120 páginasPelayoGabyZermeñoAún no hay calificaciones
- Guia Salud de Suelos Un Manual para CuidDocumento168 páginasGuia Salud de Suelos Un Manual para CuidAidee Vanessa Escobar RojasAún no hay calificaciones
- FV 28101Documento53 páginasFV 28101Juan Pablo Cubillos RomeroAún no hay calificaciones
- Figueroa 1999 PDFDocumento20 páginasFigueroa 1999 PDFDiana chaurara paradaAún no hay calificaciones
- T 1341 PDFDocumento102 páginasT 1341 PDFJuan José Aparicio PorresAún no hay calificaciones
- Digestibilidad in Vitro de NopalDocumento54 páginasDigestibilidad in Vitro de NopalprincipelicAún no hay calificaciones
- 4to Labo de Micro 2Documento59 páginas4to Labo de Micro 2jhonAún no hay calificaciones
- Universidad Nacional Agraria de La SelvaDocumento97 páginasUniversidad Nacional Agraria de La SelvaPiero GonzalesAún no hay calificaciones
- Manejo Técnico para La Producción de Aguacate Hass en La Meseta de Popayán PDFDocumento65 páginasManejo Técnico para La Producción de Aguacate Hass en La Meseta de Popayán PDFHéctor Quiñones RossAún no hay calificaciones
- Proyecto Formativo de MalezasDocumento56 páginasProyecto Formativo de MalezaswilderAún no hay calificaciones
- Oxigenoterapia en Pequeñas Especies - Universidad VeracruzanaDocumento54 páginasOxigenoterapia en Pequeñas Especies - Universidad VeracruzanaRoxana RiosAún no hay calificaciones
- Sergio Tesis Titulo 2016Documento63 páginasSergio Tesis Titulo 2016Samir Rios SacramentoAún no hay calificaciones
- Enraizante PDFDocumento112 páginasEnraizante PDFCLAUDIA YURANY ANDRADE ESCARLANTEAún no hay calificaciones
- Tesis - Evaluación de Los Parámetros Productivos Bajo 5 de SincronizaciónDocumento83 páginasTesis - Evaluación de Los Parámetros Productivos Bajo 5 de Sincronizaciónsegundo dario rios astoAún no hay calificaciones
- Anatomía Del Estómago de La Alpaca PDFDocumento33 páginasAnatomía Del Estómago de La Alpaca PDFJesús Enrique Turín VilcaAún no hay calificaciones
- Jimenez VV PDFDocumento107 páginasJimenez VV PDFDavid Ruben FigueroaAún no hay calificaciones
- Ecología y Conocimiento Tradicional de Ambystoma Altamirani - revVAA - Octubre2020 - Sin AgradecDocumento88 páginasEcología y Conocimiento Tradicional de Ambystoma Altamirani - revVAA - Octubre2020 - Sin AgradecANGELA FABIOLA CASTRO GARCIAAún no hay calificaciones
- Universidad Mayor de San Andrés: Facultad de AgronomíaDocumento153 páginasUniversidad Mayor de San Andrés: Facultad de AgronomíaDAXERAún no hay calificaciones
- T 1747Documento134 páginasT 1747angel100% (1)
- El Concepto de Degeneración en BuffonDocumento127 páginasEl Concepto de Degeneración en BuffonvioletareAún no hay calificaciones
- Guía de Los Anfibios y Reptiles Del Estado de QuerétaroDocumento145 páginasGuía de Los Anfibios y Reptiles Del Estado de QuerétaroShelby Bond100% (1)
- 2020 MonogIBADER MY-1368Documento64 páginas2020 MonogIBADER MY-1368MELISSA DARLING ORELLANA DIAZAún no hay calificaciones
- Aprendizaje del examen clínico de los equinos, bovinos y caninosDe EverandAprendizaje del examen clínico de los equinos, bovinos y caninosCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Reporte Diseccion de OjoDocumento8 páginasReporte Diseccion de OjolaputamadreAún no hay calificaciones
- Pares CranealesDocumento4 páginasPares CranealesLILIANA MONSERRAT GARCIA MALVAEZAún no hay calificaciones
- Animorphs 10 EL ANDROIDEDocumento84 páginasAnimorphs 10 EL ANDROIDEMiriam RodasAún no hay calificaciones
- Aparato Respiratorio 2020 2 (Autoguardado)Documento219 páginasAparato Respiratorio 2020 2 (Autoguardado)nicoll morenoAún no hay calificaciones
- Semana 09 Desarrollo Del CráneoDocumento34 páginasSemana 09 Desarrollo Del CráneoBambi LuAún no hay calificaciones
- Diccionario Básico de LogopediaDocumento23 páginasDiccionario Básico de LogopediaUxue EchegoyenAún no hay calificaciones
- Anatomia e Histología de Cara y Cavidad OralDocumento28 páginasAnatomia e Histología de Cara y Cavidad OralGersonAún no hay calificaciones
- Hueso Etmoides-Anatomia Cabeza & Cuello-Benites Escalante Pedro ValentinDocumento6 páginasHueso Etmoides-Anatomia Cabeza & Cuello-Benites Escalante Pedro ValentinPedro BenitesAún no hay calificaciones
- Face Yoga 5poses SpanishDocumento10 páginasFace Yoga 5poses SpanishVictoria ValenteAún no hay calificaciones
- Neurologia y Neurocirugia - 1er ParcialDocumento152 páginasNeurologia y Neurocirugia - 1er ParcialLara CazeliAún no hay calificaciones
- Exploración Radiográfica de Cráneo y Huesos Craneales.Documento20 páginasExploración Radiográfica de Cráneo y Huesos Craneales.Julio Cesar ACAún no hay calificaciones
- Periodo Pre-EmbrionarioDocumento40 páginasPeriodo Pre-EmbrionarioVeronica GonzalezAún no hay calificaciones
- CEREBELODocumento15 páginasCEREBELOMaia CucchianiAún no hay calificaciones
- Banco de Preguntas de Anatomia e Histologia 2022Documento10 páginasBanco de Preguntas de Anatomia e Histologia 2022Erick albaro zeladaAún no hay calificaciones
- Astigmatismo y PresbiciaDocumento1 páginaAstigmatismo y PresbiciaKarolyn GonzálezAún no hay calificaciones
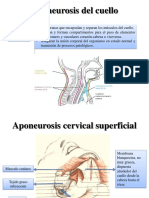
- Aponeurosis Cervical SUPERFICIAL Media y ProfundaDocumento22 páginasAponeurosis Cervical SUPERFICIAL Media y ProfundaTifany YanezAún no hay calificaciones
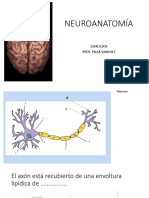
- Ejercicios de NeuroanatomíaDocumento35 páginasEjercicios de NeuroanatomíaCarmen CandilAún no hay calificaciones
- Anatomía de Fosa PosteriorDocumento44 páginasAnatomía de Fosa PosteriorTatiana FuenmayorAún no hay calificaciones
- Funciones Del Tronco EncefalicoDocumento4 páginasFunciones Del Tronco EncefalicoJesus MaresAún no hay calificaciones
- Caso Clinico CerebeloDocumento8 páginasCaso Clinico CerebeloM.465rAún no hay calificaciones
- Tesis Documento Completo FINALDocumento55 páginasTesis Documento Completo FINALDELIAAún no hay calificaciones
- Primera Mesa OftamologyDocumento7 páginasPrimera Mesa OftamologyOscar Gill VargasAún no hay calificaciones
- Analisis Cefalometrico de RickettsDocumento9 páginasAnalisis Cefalometrico de Rickettsceci hdeezAún no hay calificaciones
- Presentacion Indice de Carrea 9Documento24 páginasPresentacion Indice de Carrea 9Adriana Sanabria MendezAún no hay calificaciones
- Radiografías Extraorales. Grupo 1Documento14 páginasRadiografías Extraorales. Grupo 1Yoselin EspinAún no hay calificaciones
- Cap 1 Introducción Al Estudio Del Dolor OrofacialDocumento14 páginasCap 1 Introducción Al Estudio Del Dolor OrofacialDiana EscobarAún no hay calificaciones
- Act 5 CuidadoDocumento3 páginasAct 5 CuidadoEl más trollAún no hay calificaciones
- Nariz y Fosas NasalesDocumento4 páginasNariz y Fosas Nasalespaola balderasAún no hay calificaciones
- Respuestas Del Banco de Preguntas Cabeza y CuelloDocumento77 páginasRespuestas Del Banco de Preguntas Cabeza y CuelloCesar Orlando Torres100% (1)
- Manual de Trazado Cefalométrico de Steiner ModificadoDocumento75 páginasManual de Trazado Cefalométrico de Steiner ModificadoBruno Santiago Luna FlorezAún no hay calificaciones