También podría gustarte
- 12 Notula 77Documento15 páginas12 Notula 77juanemanuelparraAún no hay calificaciones
- GatosSocorro Libro CIBNOR 1994Documento12 páginasGatosSocorro Libro CIBNOR 1994DanielaAún no hay calificaciones
- 92173-Text de L'article-168923-1-10-20081229Documento11 páginas92173-Text de L'article-168923-1-10-20081229Mary Amore LancollAún no hay calificaciones
- Catalogo de Moluscos Litorales de ChileDocumento70 páginasCatalogo de Moluscos Litorales de ChileEdgar Bracamonte LévanoAún no hay calificaciones
- Fauna Laciana 2010 PDFDocumento25 páginasFauna Laciana 2010 PDFLydia BullockAún no hay calificaciones
- Libro Bio t2 039Documento14 páginasLibro Bio t2 039Ymir RamosAún no hay calificaciones
- Catalogo Descriptivo de Los Moluscos Litorales de Antofagasta PDFDocumento74 páginasCatalogo Descriptivo de Los Moluscos Litorales de Antofagasta PDFamendivil94Aún no hay calificaciones
- IMBERTI. 2005. Distribución Otoñal de Aves Marinas y Terrestres en Los Canales ChilenosDocumento10 páginasIMBERTI. 2005. Distribución Otoñal de Aves Marinas y Terrestres en Los Canales ChilenosPaulina Martínez CórdovaAún no hay calificaciones
- Fenología Migratoriay Movimientos Primaverales de La Codorniz Común Coturnix Coturnix en Las Islas BalearesDocumento10 páginasFenología Migratoriay Movimientos Primaverales de La Codorniz Común Coturnix Coturnix en Las Islas BalearesOtistarda2022Aún no hay calificaciones
- Registro de Los Coleopteros Acuaticos en La Región Cusco, PerúDocumento15 páginasRegistro de Los Coleopteros Acuaticos en La Región Cusco, PerúAbdhiel Arnaldo Bustamante NavarreteAún no hay calificaciones
- Hermida Et Al 20181Documento13 páginasHermida Et Al 20181CtorvalAún no hay calificaciones
- (2009) de La Ossa-V. Et Al (Pseudis Paradoxa)Documento9 páginas(2009) de La Ossa-V. Et Al (Pseudis Paradoxa)Wendy Zabala MonterrozaAún no hay calificaciones
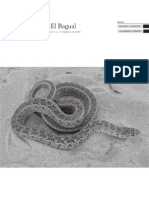
- 08 Scrocchi G J y Giraudo A R 2005 Reptiles de La Reserva El Bagual, FormosaDocumento43 páginas08 Scrocchi G J y Giraudo A R 2005 Reptiles de La Reserva El Bagual, FormosaFundacionLillo100% (7)
- Libélulas y caballitos del diablo del departamento del Meta, ColombiaDe EverandLibélulas y caballitos del diablo del departamento del Meta, ColombiaAún no hay calificaciones
- Evaluacion Lagartos AtacamaDocumento18 páginasEvaluacion Lagartos AtacamaGabriel Eduardo Mendoza RiquelmeAún no hay calificaciones
- Adami y Gordillo 1999 - Biota Asociada Macro Cyst Is PyriferaDocumento9 páginasAdami y Gordillo 1999 - Biota Asociada Macro Cyst Is PyriferaMarcelo Mayorga RodríguezAún no hay calificaciones
- Mafis: Morfología de Los Otolitos Sagitta, Lapillus y Asteriscus de Genidens Barbus de Distribución PatagónicaDocumento8 páginasMafis: Morfología de Los Otolitos Sagitta, Lapillus y Asteriscus de Genidens Barbus de Distribución PatagónicaKaren salinas araosAún no hay calificaciones
- 21 18Documento94 páginas21 18DoomwillAún no hay calificaciones
- Otaka 3Documento101 páginasOtaka 3info@ian-ani.orgAún no hay calificaciones
- 813Documento50 páginas813christina8reales8ca8Aún no hay calificaciones
- NUMEROSAS PAREJAS DE AGUILUCHO COMÚN (Buteo Polyosoma) NIDIFICANDO EN POSTES DE ELECTRICIDAD EN EL NORTE PATAGÓNICO, ARGENTINADocumento10 páginasNUMEROSAS PAREJAS DE AGUILUCHO COMÚN (Buteo Polyosoma) NIDIFICANDO EN POSTES DE ELECTRICIDAD EN EL NORTE PATAGÓNICO, ARGENTINASara JohnsonAún no hay calificaciones
- 2019barragan VazquezetalReptilesDocumento11 páginas2019barragan VazquezetalReptilesLuz Ariadna Gallardo RecinoAún no hay calificaciones
- Naturalmente Public Ac in IndividualDocumento2 páginasNaturalmente Public Ac in IndividualALEJANDRO OSORIO GÓMEZAún no hay calificaciones
- BosqueDocumento8 páginasBosqueNorberto MuzzachiodiAún no hay calificaciones
- Diversity of Scorpions (Chelicerata) in The Department of Atlántico, ColombiaDocumento15 páginasDiversity of Scorpions (Chelicerata) in The Department of Atlántico, ColombiaMiguel GutierrezAún no hay calificaciones
- Cuenca Bescoset1996MMVallipnDocumento34 páginasCuenca Bescoset1996MMVallipnDanna Brigith Vera CarrilloAún no hay calificaciones
- DytiscidaeDocumento5 páginasDytiscidaeGis CAún no hay calificaciones
- Valladaresetal 2021 LbasadreiDocumento14 páginasValladaresetal 2021 Lbasadreiruthmcontreras88Aún no hay calificaciones
- Diversidad de RotiferosDocumento10 páginasDiversidad de Rotiferosfrancisco_jycAún no hay calificaciones
- Rastros de Murciélagos PDFDocumento10 páginasRastros de Murciélagos PDFanzaeduardo1Aún no hay calificaciones
- Guia de Campo de Los Mamiferos Del EcuadDocumento3 páginasGuia de Campo de Los Mamiferos Del EcuadNutriRit EssentialAún no hay calificaciones
- Gasteropodos Marinos ColumbretesDocumento39 páginasGasteropodos Marinos ColumbretesPerico PalotesAún no hay calificaciones
- Historia Natural 2017 1 Art 45 571Documento14 páginasHistoria Natural 2017 1 Art 45 57186 leonelliniAún no hay calificaciones
- 1 PBDocumento18 páginas1 PBAlexander LopezteguiAún no hay calificaciones
- 10 Notula 103Documento10 páginas10 Notula 103juanemanuelparraAún no hay calificaciones
- COCUYO. Revista Cubana de ZoologiaDocumento33 páginasCOCUYO. Revista Cubana de ZoologiaAndresr Ramírez100% (1)
- Elmidae Del NeotropicoDocumento14 páginasElmidae Del Neotropicoadbs91Aún no hay calificaciones
- RPB - 1999!6!1!5!47 Estado Actual Del Conocimiento de Los Moluscos Acuáticos en El Perú Paredes Et AlDocumento45 páginasRPB - 1999!6!1!5!47 Estado Actual Del Conocimiento de Los Moluscos Acuáticos en El Perú Paredes Et AlAracelly ChavezAún no hay calificaciones
- Murcielagosdelaciudadde MexicoDocumento26 páginasMurcielagosdelaciudadde MexicoMartín GutiérrezAún no hay calificaciones
- Biología de Las Moscas CalliphoridaeDocumento8 páginasBiología de Las Moscas CalliphoridaeRafael ÁvilaAún no hay calificaciones
- 01 Chocolate L 2Documento6 páginas01 Chocolate L 2perezyajaira958Aún no hay calificaciones
- Reproduccinymadurezgondicadelalisa MugilDocumento15 páginasReproduccinymadurezgondicadelalisa MugilAldris Anuar Geliz DiazAún no hay calificaciones
- Dialnet NotasCorologicasParaLaFloraDeMallorcaIII 9028838Documento14 páginasDialnet NotasCorologicasParaLaFloraDeMallorcaIII 9028838jlccAún no hay calificaciones
- Aves Rapaces Como Indicadoras Ambientales en La Jurisdicción de La CARDocumento17 páginasAves Rapaces Como Indicadoras Ambientales en La Jurisdicción de La CARJennifer Ardila AyalaAún no hay calificaciones
- Insect Oster Rest ResDocumento32 páginasInsect Oster Rest ResCandela GaunaAún no hay calificaciones
- Instituto Tecnologico de MexicoDocumento28 páginasInstituto Tecnologico de MexicolelyAún no hay calificaciones
- Ecuador-Peces de CuyabenoDocumento55 páginasEcuador-Peces de CuyabenoRuth Zambrano MonteroAún no hay calificaciones
- Avendaño Et Al., 1998Documento7 páginasAvendaño Et Al., 1998JHORDY ADISONSCOT ARMAS AMATAún no hay calificaciones
- 473-Texto Del Artículo-503-1-10-20140122Documento12 páginas473-Texto Del Artículo-503-1-10-20140122Marcelino BertranAún no hay calificaciones
- Cladoceros Colombia PDFDocumento42 páginasCladoceros Colombia PDFroraysi_cm01Aún no hay calificaciones
- Anadara Repro Recluta (2004 185)Documento11 páginasAnadara Repro Recluta (2004 185)Ronny GongoraAún no hay calificaciones
- INVESTIGACIÓN Y CONSERVACIÓN SOBRE MURCIÉLAGOS EN EL ECUADORDe EverandINVESTIGACIÓN Y CONSERVACIÓN SOBRE MURCIÉLAGOS EN EL ECUADORAún no hay calificaciones
- Grismado 2007 Arañas de OtamendiDocumento95 páginasGrismado 2007 Arañas de OtamendiGabriel Maugeri100% (1)
- Zoologia de Invertebrados 2Documento33 páginasZoologia de Invertebrados 2AlejanDrah BarriiOsAún no hay calificaciones
- Dieta Del Biguá (Phalacrocorax Olivaceus) Durante La Primavera en El Estuario de Bahía Blanca, Buenos Aires, ArgentinaDocumento6 páginasDieta Del Biguá (Phalacrocorax Olivaceus) Durante La Primavera en El Estuario de Bahía Blanca, Buenos Aires, ArgentinaTalita BackhausAún no hay calificaciones
- Informe N°7 ReptilesDocumento15 páginasInforme N°7 ReptilesJhom AlquichireAún no hay calificaciones
- Avifauna neotropical : ecología y conservaciónDe EverandAvifauna neotropical : ecología y conservaciónAún no hay calificaciones
- Temolines. Los coleópteros entre los antiguos mexicanosDe EverandTemolines. Los coleópteros entre los antiguos mexicanosAún no hay calificaciones
- Catálogo de los copépodos (Crustacea: Copepoda: Calanoida y Cyclopoida) de cuerpos de agua temporales de Jalisco, México: Taxonomía y distribuciónDe EverandCatálogo de los copépodos (Crustacea: Copepoda: Calanoida y Cyclopoida) de cuerpos de agua temporales de Jalisco, México: Taxonomía y distribuciónAún no hay calificaciones
- Ecología y conservación en los mares de MéxicoDe EverandEcología y conservación en los mares de MéxicoAún no hay calificaciones
- Crucigramas - Biomoleculas OrgánicasDocumento4 páginasCrucigramas - Biomoleculas OrgánicasLuisa GonzalezAún no hay calificaciones
- Metabolismo Del Cáncer Mirando Hacia El Futuro (1) EspañolDocumento12 páginasMetabolismo Del Cáncer Mirando Hacia El Futuro (1) EspañolJOSE ANTONIO GOMEZ LA ROSAAún no hay calificaciones
- BIOLOGIADocumento2 páginasBIOLOGIACarlos PerezAún no hay calificaciones
- Folleto Del MaizDocumento2 páginasFolleto Del MaizCatalina100% (1)
- Anemia Sideroblastica Epidemiologia Novedades ClinicaDocumento9 páginasAnemia Sideroblastica Epidemiologia Novedades ClinicaItzel LoesAún no hay calificaciones
- Practica 9Documento10 páginasPractica 9CELIA SALCEDO ESCOBARAún no hay calificaciones
- Práctica N°07 - Bma - LervDocumento3 páginasPráctica N°07 - Bma - LervJhiampier adicionalAún no hay calificaciones
- Dcm134o PDFDocumento2 páginasDcm134o PDFandrostona980Aún no hay calificaciones
- Accidentes Ofídicos en Pequeñas Especies Caninos y FelinosDocumento16 páginasAccidentes Ofídicos en Pequeñas Especies Caninos y FelinosERIKA ARCE GAVIRIAAún no hay calificaciones
- Informe Practico de BiologíaDocumento6 páginasInforme Practico de BiologíaMelchorita stephanie Nuñez CordovaAún no hay calificaciones
- Enzimas+Especiales+Granzimas+y+Abzimas+ +grupo+4Documento33 páginasEnzimas+Especiales+Granzimas+y+Abzimas+ +grupo+4Anypsa Palacios RomanAún no hay calificaciones
- Glosario - Ciclo CelularDocumento4 páginasGlosario - Ciclo CelularDaniela100% (2)
- Triptico Reino ProtoctistaDocumento2 páginasTriptico Reino ProtoctistaLaura GarciaAún no hay calificaciones
- Ejercicios Seleccion2025Documento7 páginasEjercicios Seleccion2025Junior FloresAún no hay calificaciones
- Los Genes y El AdnDocumento8 páginasLos Genes y El Adnhercilia PerezAún no hay calificaciones
- Clase 7 Reparacion Del DNADocumento13 páginasClase 7 Reparacion Del DNAKatya JaramilloAún no hay calificaciones
- Segunda Tarea EticaDocumento10 páginasSegunda Tarea EticaluisAún no hay calificaciones
- Célula y Su ImportanciaDocumento3 páginasCélula y Su Importanciacandy martinezAún no hay calificaciones
- Actividad 2 - Ud4 BiomolDocumento3 páginasActividad 2 - Ud4 BiomolnelooxdAún no hay calificaciones
- Tarea 3 - Identificación Morfológica y Organográfica de Las PlantasDocumento14 páginasTarea 3 - Identificación Morfológica y Organográfica de Las PlantasAstrid Carolina Leal AlvarezAún no hay calificaciones
- Evaluación 1B - Guardianes de Nuestros HermanosDocumento5 páginasEvaluación 1B - Guardianes de Nuestros HermanosJanet ParraAún no hay calificaciones
- Salud PublicaDocumento5 páginasSalud PublicaABRIL NAOMI BOCK GARCIAAún no hay calificaciones
- Ingeniería Civil en Bioinformática: Plan de EstudiosDocumento1 páginaIngeniería Civil en Bioinformática: Plan de EstudiosIgnacio MoralesAún no hay calificaciones
- Trabajo Sem 2 LeucemiaDocumento14 páginasTrabajo Sem 2 LeucemiaEurisAún no hay calificaciones
- Hongos y LevadurasDocumento38 páginasHongos y LevadurasAstrid MartinezAún no hay calificaciones
- Tarea # 1 Evaluacion de La InteligenciaDocumento8 páginasTarea # 1 Evaluacion de La Inteligenciayaniris jaquezAún no hay calificaciones
- Biomoleculas y Su Interacción en La CélulaDocumento53 páginasBiomoleculas y Su Interacción en La CélulaIvan MontañoAún no hay calificaciones
- CELULADocumento6 páginasCELULAalejandra rAún no hay calificaciones
- Ejercicios CladísticaDocumento22 páginasEjercicios CladísticaYensy AmadoAún no hay calificaciones
- MBSMT1Documento57 páginasMBSMT1Gerdel alberto cantillo barriosAún no hay calificaciones