También podría gustarte
- Hoja de Reporte de Laboratorio Nº3 Curvas V-IDocumento12 páginasHoja de Reporte de Laboratorio Nº3 Curvas V-IFulano Mengano Sutano SerranoAún no hay calificaciones
- Hoja de Reporte de Laboratorio Nº3 Curvas V-IDocumento12 páginasHoja de Reporte de Laboratorio Nº3 Curvas V-IFulano Mengano Sutano SerranoAún no hay calificaciones
- Un Libro, Tiempo y CaosDocumento1 páginaUn Libro, Tiempo y CaosFulano Mengano Sutano SerranoAún no hay calificaciones
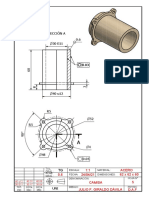
- CAMISADocumento1 páginaCAMISAFulano Mengano Sutano SerranoAún no hay calificaciones
- Ensayo de Traccion - MC112-2021-1Documento10 páginasEnsayo de Traccion - MC112-2021-1Fulano Mengano Sutano SerranoAún no hay calificaciones
- Sección C-PC2 FIII 2020-2 VidalDocumento10 páginasSección C-PC2 FIII 2020-2 VidalFulano Mengano Sutano SerranoAún no hay calificaciones
- Pc2 Fisiii 2021-2Documento5 páginasPc2 Fisiii 2021-2Fulano Mengano Sutano SerranoAún no hay calificaciones
- Ensayo de Traccion - MC112-2021-1Documento11 páginasEnsayo de Traccion - MC112-2021-1Fulano Mengano Sutano SerranoAún no hay calificaciones
- Ejercicio Ajustes JZCDocumento3 páginasEjercicio Ajustes JZCFulano Mengano Sutano SerranoAún no hay calificaciones
- Nota Del Examen (1Documento4 páginasNota Del Examen (1Zedrix Augusto Quispe CarrilloAún no hay calificaciones
- PETRODocumento2 páginasPETROFulano Mengano Sutano SerranoAún no hay calificaciones
- UbiDocumento1 páginaUbiFulano Mengano Sutano SerranoAún no hay calificaciones
- Ensayo de Traccion - MC112-2021-1Documento11 páginasEnsayo de Traccion - MC112-2021-1Fulano Mengano Sutano SerranoAún no hay calificaciones
- TuberiasDocumento90 páginasTuberiasFulano Mengano Sutano Serrano100% (2)
- Expo Del DiaDocumento5 páginasExpo Del DiaFulano Mengano Sutano SerranoAún no hay calificaciones
- Seminario ADocumento22 páginasSeminario AFulano Mengano Sutano SerranoAún no hay calificaciones
- SeccC-PC1 FISIII 2020-2Documento6 páginasSeccC-PC1 FISIII 2020-2Fulano Mengano Sutano SerranoAún no hay calificaciones
- SILABO-BQU01-Química I-2020-31-05-20Documento4 páginasSILABO-BQU01-Química I-2020-31-05-20Pablo Cesar VadilloAún no hay calificaciones
- Momento CentralDocumento17 páginasMomento CentralFulano Mengano Sutano SerranoAún no hay calificaciones
- Seminario BDocumento18 páginasSeminario BFulano Mengano Sutano SerranoAún no hay calificaciones
- Prueba de Entrada 2021-2Documento1 páginaPrueba de Entrada 2021-2Fulano Mengano Sutano SerranoAún no hay calificaciones
- 2 Doc 2 Solución Es MB148 2021 IDocumento6 páginas2 Doc 2 Solución Es MB148 2021 IFulano Mengano Sutano SerranoAún no hay calificaciones
- PROBLEMA 03. - (7 Puntos)Documento1 páginaPROBLEMA 03. - (7 Puntos)Fulano Mengano Sutano SerranoAún no hay calificaciones
- Sustitutorio MB545 2021 IDocumento3 páginasSustitutorio MB545 2021 IFulano Mengano Sutano SerranoAún no hay calificaciones
- Sustitutorio MB545 2021 IDocumento3 páginasSustitutorio MB545 2021 IFulano Mengano Sutano SerranoAún no hay calificaciones
- 1 Doc 1 Preguntas Es MB148 2021 IDocumento1 página1 Doc 1 Preguntas Es MB148 2021 IFulano Mengano Sutano SerranoAún no hay calificaciones
- Mc509 Examen Sustitutorio 2021-1 IndicacionesDocumento1 páginaMc509 Examen Sustitutorio 2021-1 IndicacionesFulano Mengano Sutano SerranoAún no hay calificaciones
- PROBLEMA 02. - (7 Puntos)Documento1 páginaPROBLEMA 02. - (7 Puntos)Fulano Mengano Sutano SerranoAún no hay calificaciones
- Art 14Documento4 páginasArt 14Jonathan GutierrezAún no hay calificaciones
- Circulatorio AnimalDocumento28 páginasCirculatorio Animalgato sexoAún no hay calificaciones
- Partes de Una Antena de InsectosDocumento8 páginasPartes de Una Antena de InsectosZeyNep Cl DaraAún no hay calificaciones
- Oso PandaDocumento8 páginasOso PandaEvelinChingayTorresAún no hay calificaciones
- 13 Animales Que Podrían Extinguirse Este 2015Documento2 páginas13 Animales Que Podrían Extinguirse Este 2015MILCA LARAAún no hay calificaciones
- Actividades NuevasDocumento7 páginasActividades NuevasMelina Ivana BragaAún no hay calificaciones
- RA-RAU-SSO-PUT-PRO-15 Procedimiento de Uso y Traslado Del Auto-Rescatador Gokang K-S30-OkDocumento4 páginasRA-RAU-SSO-PUT-PRO-15 Procedimiento de Uso y Traslado Del Auto-Rescatador Gokang K-S30-OkLuis FrancoAún no hay calificaciones
- TripticoDocumento2 páginasTripticoyueshtisuw42Aún no hay calificaciones
- Web Mapa BIOPARC Valencia Julio 2020Documento1 páginaWeb Mapa BIOPARC Valencia Julio 2020VivoyversoAún no hay calificaciones
- 4a Práctica Lab Anatomia Del Cuello 2a ParteDocumento4 páginas4a Práctica Lab Anatomia Del Cuello 2a ParteRuth Cobo RosalesAún no hay calificaciones
- Animales AlbunDocumento15 páginasAnimales AlbunRojas Santiago LuisAún no hay calificaciones
- Reino Animal ActividadesDocumento5 páginasReino Animal ActividadesElenaAún no hay calificaciones
- FICHA - SARDINA - 1º de ESODocumento2 páginasFICHA - SARDINA - 1º de ESOAlejandro RomeroAún no hay calificaciones
- CapibaraDocumento9 páginasCapibaraDaniAún no hay calificaciones
- HORMIGASDocumento4 páginasHORMIGASDenisse Sosa OrtizAún no hay calificaciones
- AnfibiosDocumento74 páginasAnfibiosMiguel SanchezAún no hay calificaciones
- Taxonomía Del PerroDocumento7 páginasTaxonomía Del PerroMaria Ximena Calderon Vaca0% (1)
- Untitled 2Documento13 páginasUntitled 2Darwin Hidalgo Suni SuniAún no hay calificaciones
- Hominización - InfografiaDocumento3 páginasHominización - InfografiaSantiago Pavajeau ReyesAún no hay calificaciones
- La Viuda NegraDocumento2 páginasLa Viuda NegraNeri GarciaAún no hay calificaciones
- Cnidarios Dulceacuicola: Las HidrasDocumento8 páginasCnidarios Dulceacuicola: Las HidrasDiego MyxomycotaAún no hay calificaciones
- Puente ComunerosDocumento15 páginasPuente ComunerosMessias ScrAún no hay calificaciones
- Ácaros de La Sarna - CVDocumento34 páginasÁcaros de La Sarna - CVErwin AyalaAún no hay calificaciones
- Unicelulares y PluricelularesDocumento2 páginasUnicelulares y PluricelularesTao-Tao ʕ •ᴥ• ʔAún no hay calificaciones
- EntoproctaDocumento6 páginasEntoproctaJorobadoAún no hay calificaciones
- Peces Definitivo.Documento15 páginasPeces Definitivo.Christopher Andre Cavero RiveroAún no hay calificaciones
- Seres Vivos La Diversidad 2014Documento20 páginasSeres Vivos La Diversidad 2014jonvelAún no hay calificaciones
- 30 Fábulas Cortas para Niños de PrimariaDocumento40 páginas30 Fábulas Cortas para Niños de PrimariaJunior Lopez LAún no hay calificaciones
- Animales 1 2Documento6 páginasAnimales 1 2Paz Correa SuárezAún no hay calificaciones
- Anaya 2quinto Refuerzo y Ampliación de Conocimiento Del MedioDocumento72 páginasAnaya 2quinto Refuerzo y Ampliación de Conocimiento Del MedioMiriam BarbaAún no hay calificaciones
- Paso5 - Zuleima Papamija - P.Rumiantes - ReacticaDocumento11 páginasPaso5 - Zuleima Papamija - P.Rumiantes - ReacticaZuleima PapamijaAún no hay calificaciones