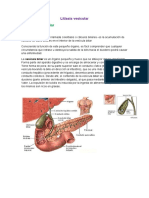
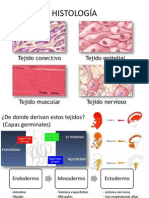
También podría gustarte
- TrofoterapiaDocumento46 páginasTrofoterapiaGerifalte89% (36)
- Calificación BDI de BeckDocumento3 páginasCalificación BDI de BeckRosalia OA100% (1)
- Desarrollo Del Sistema NerviosoDocumento6 páginasDesarrollo Del Sistema NerviosoFelipe Poblete VegaAún no hay calificaciones
- Semiologia Tercer Año USACDocumento297 páginasSemiologia Tercer Año USACHanTAún no hay calificaciones
- Embriologia Power Point Ciclo OvaricoDocumento22 páginasEmbriologia Power Point Ciclo OvaricoLivanessa Ubri GonzalezAún no hay calificaciones
- Caso Steve. Analisis Multiaxial.Documento2 páginasCaso Steve. Analisis Multiaxial.Camilo Rubiano100% (1)
- INSPECCIÓNDocumento20 páginasINSPECCIÓNalfredoAún no hay calificaciones
- Análisis Amplificado de Los ChakrasDocumento6 páginasAnálisis Amplificado de Los ChakrasNestor Fernandez RiveroAún no hay calificaciones
- UrogenitalDocumento40 páginasUrogenitalElianneAún no hay calificaciones
- ABDOMEN InspeccionDocumento3 páginasABDOMEN InspeccionSonny LoayesAún no hay calificaciones
- CorazónDocumento10 páginasCorazónJL Díaz0% (1)
- Infografia Sobre La NOM 016 SSA3 2012Documento1 páginaInfografia Sobre La NOM 016 SSA3 2012Liza Maria LopezAún no hay calificaciones
- Seminario 25 - Hemorragias ObtetricasDocumento35 páginasSeminario 25 - Hemorragias ObtetricasGisela Niebuhr Herrera100% (1)
- Importancia Del Volumen ResidualDocumento1 páginaImportancia Del Volumen ResidualStephany Conde PoloAún no hay calificaciones
- Diphyllobothrium LatumDocumento15 páginasDiphyllobothrium LatumMelissa Peña RoblesAún no hay calificaciones
- Seminario (Tetralogía de Fallot)Documento19 páginasSeminario (Tetralogía de Fallot)Angie DanielaAún no hay calificaciones
- 01.-Artritis Reumatoidea, Les, Vasculitis USS - TEORIADocumento55 páginas01.-Artritis Reumatoidea, Les, Vasculitis USS - TEORIAMaritho Vera SànchezAún no hay calificaciones
- Ensayo SuicidioDocumento27 páginasEnsayo SuicidioJosé Luis AguilarAún no hay calificaciones
- Manual de Histopatologia 13-2-1Documento56 páginasManual de Histopatologia 13-2-1Psyche Calderon VargasAún no hay calificaciones
- Tratamientos Tabla Equivalencia OpioidesDocumento6 páginasTratamientos Tabla Equivalencia OpioidesoscarAún no hay calificaciones
- MEL y Master Luong Minh DangDocumento2 páginasMEL y Master Luong Minh Dangfloralop1100% (1)
- Faros y FocosDocumento13 páginasFaros y Focosalexander mendoza50% (2)
- Desarrollo de Vellosidades CorionicasDocumento11 páginasDesarrollo de Vellosidades CorionicasAlexia AlvaradoAún no hay calificaciones
- Placenta y Membranas Fetales-Mantilla Ibañez.Documento10 páginasPlacenta y Membranas Fetales-Mantilla Ibañez.María Luisa MantillaAún no hay calificaciones
- Placenta PDFDocumento14 páginasPlacenta PDFGallardo RichardAún no hay calificaciones
- Morfogenesis de La PlacentaDocumento17 páginasMorfogenesis de La PlacentajjgenconAún no hay calificaciones
- Cap 14 y 15Documento4 páginasCap 14 y 15mechefierroAún no hay calificaciones
- Cavidades CorporalesDocumento5 páginasCavidades CorporalesYasmin JimenezAún no hay calificaciones
- Tema 13. Cavidades Corporales y SerosasDocumento8 páginasTema 13. Cavidades Corporales y SerosasVibe NationAún no hay calificaciones
- Embriologia Del Sistema RespiratorioDocumento44 páginasEmbriologia Del Sistema Respiratorioluisortega93Aún no hay calificaciones
- Placenta MacroscópicaDocumento11 páginasPlacenta MacroscópicaLiliana Julissa García AlarcónAún no hay calificaciones
- OVOGENESISDocumento29 páginasOVOGENESISGenessis Cardenas100% (1)
- Histología RiñónDocumento7 páginasHistología RiñónSneider Alexander Garcia TascoAún no hay calificaciones
- Desarrollo de Cavidades CorporalesDocumento33 páginasDesarrollo de Cavidades CorporalesDiana OrtegaAún no hay calificaciones
- Ciclo Menstrual FemeninoDocumento6 páginasCiclo Menstrual FemeninoMARCELO MIGUELAún no hay calificaciones
- Enfermedades Del Tracto GastrointestinalDocumento61 páginasEnfermedades Del Tracto GastrointestinalRodrickAún no hay calificaciones
- Tejido CartilaginosoDocumento36 páginasTejido CartilaginosoJessi Ramirez UAún no hay calificaciones
- Desarrollo Del Tubo Intestinal, Páncreas y DiafragmaDocumento6 páginasDesarrollo Del Tubo Intestinal, Páncreas y Diafragmaeliana bustamante0% (1)
- Guia IsoinmunizacionDocumento10 páginasGuia IsoinmunizacionAxl MendezAún no hay calificaciones
- Anatomía Microscópica Del Sistema CirculatorioDocumento5 páginasAnatomía Microscópica Del Sistema CirculatorioKarina BastidaAún no hay calificaciones
- Cavidades Corporales, Mesenterios y DiafragmaDocumento51 páginasCavidades Corporales, Mesenterios y DiafragmaKaren Hernàndez67% (3)
- Anomalías de Anexos EmbrionariosDocumento3 páginasAnomalías de Anexos EmbrionariosJessi Márquez100% (1)
- PlacentaDocumento29 páginasPlacentainextremus_cayado7754100% (1)
- Embriologia Del Sistema Nervioso DR Carlos Alejandro Gonzales Medina UNMSMDocumento83 páginasEmbriologia Del Sistema Nervioso DR Carlos Alejandro Gonzales Medina UNMSMCarlos GonzalesAún no hay calificaciones
- Anomalias PlacentariasDocumento85 páginasAnomalias PlacentariasLuis Enrique LaraAún no hay calificaciones
- Mapa Conceptual - Desarrollo Del CorazónDocumento1 páginaMapa Conceptual - Desarrollo Del CorazónDagmar MarquezAún no hay calificaciones
- Resuemn Del Tejido Glandular 1Documento3 páginasResuemn Del Tejido Glandular 1Silvana Branca FerrerAún no hay calificaciones
- Histología Del RiñónDocumento1 páginaHistología Del RiñónAlondra Alitzel GarciaAún no hay calificaciones
- Primera Semana de GestaciónDocumento3 páginasPrimera Semana de GestaciónAmal CalcagniAún no hay calificaciones
- Anato VejigaDocumento4 páginasAnato VejigaEmanuel MartinezAún no hay calificaciones
- Sistema Cardiovascular. EMBRIOLOGIA PDFDocumento10 páginasSistema Cardiovascular. EMBRIOLOGIA PDFDavid De La RosaAún no hay calificaciones
- Diferencias Entre Celulas Procariotas y EucariotasDocumento2 páginasDiferencias Entre Celulas Procariotas y Eucariotasmells19Aún no hay calificaciones
- Litiasis VesicularDocumento3 páginasLitiasis VesicularAngel Rual Reyes CastroAún no hay calificaciones
- Desarrollo Del Sistema TegumentarioDocumento104 páginasDesarrollo Del Sistema TegumentarioGianella VDKAún no hay calificaciones
- Auscultación CardíacaDocumento21 páginasAuscultación CardíacaAlezita AccostaAún no hay calificaciones
- Intestino Medio y PosteriorDocumento78 páginasIntestino Medio y PosteriorDayma SantamariaAún no hay calificaciones
- Embriologia Del Sistema CardiovascularDocumento13 páginasEmbriologia Del Sistema CardiovascularMirgelys VargasAún no hay calificaciones
- Tercera y Octava Semana de Desarrollo EmbrionarioDocumento6 páginasTercera y Octava Semana de Desarrollo EmbrionarioXiomara LuengoAún no hay calificaciones
- Asfixia Perinatal.Documento21 páginasAsfixia Perinatal.German Andres ZambranoAún no hay calificaciones
- Embriología Del Aparato Reproductor FemeninoDocumento9 páginasEmbriología Del Aparato Reproductor FemeninoAlberty RodriguezAún no hay calificaciones
- HISTOLOGÍADocumento55 páginasHISTOLOGÍAConstanza PalavecinoAún no hay calificaciones
- El Tubo Intestinal y Las Cavidades CorporalesDocumento48 páginasEl Tubo Intestinal y Las Cavidades CorporalesUlises García Arellano100% (7)
- Membranas Fetales y PlacentaDocumento15 páginasMembranas Fetales y PlacentaGuillermo Gaona BojorquezAún no hay calificaciones
- Movimiento Celular FISIOLOGÍA (Guyton)Documento11 páginasMovimiento Celular FISIOLOGÍA (Guyton)GeralKatiaCruzYufraAún no hay calificaciones
- Formación y Desarrollo de La PlacentaDocumento2 páginasFormación y Desarrollo de La PlacentaYanela AlfaroAún no hay calificaciones
- Características Morfológicas de La PlacentaDocumento2 páginasCaracterísticas Morfológicas de La PlacentaCarlos Aguilar MejíaAún no hay calificaciones
- Cambios Placentarios Al Final Del Embarazo InformeDocumento10 páginasCambios Placentarios Al Final Del Embarazo Informelm8322074Aún no hay calificaciones
- 1.2 Anexos Fetales y Líquido AmnióticoDocumento16 páginas1.2 Anexos Fetales y Líquido AmnióticoPao VasconezAún no hay calificaciones
- Desarrollo y Funcion de Los Anexos OvularesDocumento89 páginasDesarrollo y Funcion de Los Anexos OvularesRamses Santos0% (1)
- 1 Placenta y Anexos OvularesDocumento40 páginas1 Placenta y Anexos OvularesClelia LoyolaAún no hay calificaciones
- Efectos Biológicos de La MarihuanaDocumento15 páginasEfectos Biológicos de La MarihuanaAlejandro Ordóñez25% (4)
- CONSENTIMIENTO FX Tibia y EroneDocumento2 páginasCONSENTIMIENTO FX Tibia y EroneAndrés ToscanoAún no hay calificaciones
- Anexo 10 FICHA DE LIMPIEZA Y DESINFECCION NUEVODocumento2 páginasAnexo 10 FICHA DE LIMPIEZA Y DESINFECCION NUEVOElmer OreatsAún no hay calificaciones
- Valdemar Rojas, Apuntes para Una BiografíaDocumento143 páginasValdemar Rojas, Apuntes para Una BiografíaSarelly Martínez Mendoza100% (1)
- Lectura y Escritura en Baja VisiónDocumento16 páginasLectura y Escritura en Baja VisiónArturo SuncinAún no hay calificaciones
- Eunacom 2013 PreguntasDocumento14 páginasEunacom 2013 Preguntaselcaliche80% (5)
- Mejor Espalda Mejor CuerpoDocumento90 páginasMejor Espalda Mejor CuerporvegafAún no hay calificaciones
- Infecciones Del Tracto Urinario en Bebés Mayores de Un Mes y Niños Menores de Dos AñosDocumento15 páginasInfecciones Del Tracto Urinario en Bebés Mayores de Un Mes y Niños Menores de Dos AñoscanahicordovasalazarAún no hay calificaciones
- Historia de Los ArticuladoresDocumento27 páginasHistoria de Los ArticuladoresHugo CeliAún no hay calificaciones
- Plasmodiophora BrassicaeDocumento4 páginasPlasmodiophora BrassicaeMarco BermudezAún no hay calificaciones
- Ejercicios Modelo 1 y 2Documento4 páginasEjercicios Modelo 1 y 2Xavier Enrique SanchezAún no hay calificaciones
- LibrodesdeomnipageDocumento199 páginasLibrodesdeomnipagealbertoriosvAún no hay calificaciones
- Consentimiento Laser OnicomicosisDocumento2 páginasConsentimiento Laser Onicomicosismedspa.casa.alma.divinaAún no hay calificaciones
- AO y AAOSDocumento4 páginasAO y AAOSCesarIslas100% (1)
- TECNICA Conjunto de Saberes Prácticos o Procedimientos para Obtener Un ResultadoDocumento11 páginasTECNICA Conjunto de Saberes Prácticos o Procedimientos para Obtener Un ResultadoShama Hip-Nosis75% (4)
- 14 Flujogramas Laboratorios PDFDocumento9 páginas14 Flujogramas Laboratorios PDFAlan Nelson Siles ArandiaAún no hay calificaciones
- Diagnóstico Diferencial de Ojo Rojo PUCDocumento12 páginasDiagnóstico Diferencial de Ojo Rojo PUCMichael SerranoAún no hay calificaciones
- Cloruro de CalcioDocumento18 páginasCloruro de CalcioERNICHE GUEVARAAún no hay calificaciones
- Ficha de Atencion Podológica 23.07.2023Documento5 páginasFicha de Atencion Podológica 23.07.2023alu.alonsoibanez.cachAún no hay calificaciones
- Clase 3Documento56 páginasClase 3AntonellaAún no hay calificaciones