También podría gustarte
- Shock PBDocumento36 páginasShock PBPaula Andrea Beltran Ramirez100% (1)
- Los Servicios de EnfermeriaDocumento20 páginasLos Servicios de EnfermeriaANNA MARIANNE CHOLAN TARRILLOAún no hay calificaciones
- ESPIROMETRÍADocumento8 páginasESPIROMETRÍADavid QuizhpeAún no hay calificaciones
- Fisiología Del EmbarazoDocumento13 páginasFisiología Del EmbarazoKaren RivasplataAún no hay calificaciones
- Tabla 7. Preparación de Diluciones de Agentes Antimicrobianos para Usar en Pruebas de Susceptibilidad de Dilución en AgarDocumento43 páginasTabla 7. Preparación de Diluciones de Agentes Antimicrobianos para Usar en Pruebas de Susceptibilidad de Dilución en AgarSaul PalavecinoAún no hay calificaciones
- Tecnica de Vias de Administracion ParenteralDocumento5 páginasTecnica de Vias de Administracion ParenteralfridafcAún no hay calificaciones
- Fármacos AntihipertensivosDocumento62 páginasFármacos AntihipertensivosGustavo J. Miraval CastroAún no hay calificaciones
- Asistente de Selección - TelefinperDocumento41 páginasAsistente de Selección - TelefinperJohoSha La RosaAún no hay calificaciones
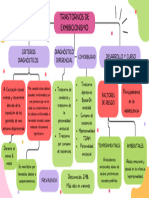
- Teorías Del EnvejecimientoDocumento9 páginasTeorías Del Envejecimientoyamilet VazquezAún no hay calificaciones
- Procedimiento de Limpieza y Desinfección de Oficinas y Zonas Comunes Limpieza y Desinfección de Las OficinasDocumento5 páginasProcedimiento de Limpieza y Desinfección de Oficinas y Zonas Comunes Limpieza y Desinfección de Las OficinasAbel Gilberto Acosta CornejoAún no hay calificaciones
- Resolución 3512 DEL 2019 PBSDocumento114 páginasResolución 3512 DEL 2019 PBSandres sotoAún no hay calificaciones
- Resumen Plan Nacional Salud Mental Adicciones.Documento5 páginasResumen Plan Nacional Salud Mental Adicciones.Montevideo PortalAún no hay calificaciones
- Ventajas y Desventajas de La Ligadura de TrompasDocumento6 páginasVentajas y Desventajas de La Ligadura de TrompasKenniaAún no hay calificaciones
- Trastornos de ExhibicionismoDocumento1 páginaTrastornos de ExhibicionismoREGINA AYOAún no hay calificaciones
- Disrafia EspinalDocumento11 páginasDisrafia EspinalHbc UrrunagaAún no hay calificaciones
- APOSTOSISDocumento26 páginasAPOSTOSISlizzy chanAún no hay calificaciones
- Ventilacià N Mecanica Septiembre 2023Documento42 páginasVentilacià N Mecanica Septiembre 2023DAN-SOCKHSC ZAún no hay calificaciones
- Anexo I-3 CLASIFICACION DE LAS PACTICAS POR CATEGORIA DE RIESGODocumento3 páginasAnexo I-3 CLASIFICACION DE LAS PACTICAS POR CATEGORIA DE RIESGOKatherine VenegasAún no hay calificaciones
- Cuestionario BioanalisisDocumento26 páginasCuestionario BioanalisisLiliana MarinAún no hay calificaciones
- Psicología Clínica Infantojuvenil - Trastornos de Síntomas SomáticosDocumento9 páginasPsicología Clínica Infantojuvenil - Trastornos de Síntomas SomáticosJohanna Quintero MarulandaAún no hay calificaciones
- Patología Corazón y TiroidesDocumento90 páginasPatología Corazón y TiroidesLuis Miguel Melendez SanchezAún no hay calificaciones
- Modelos de EnfermedadDocumento21 páginasModelos de EnfermedadSnorreteAún no hay calificaciones
- Infecciones NosocomialesDocumento7 páginasInfecciones Nosocomiales506070Aún no hay calificaciones
- PB Assist Plus Pip SPDocumento4 páginasPB Assist Plus Pip SPFrancisco VegasAún no hay calificaciones
- Enterobius Vermicularis-2011Documento16 páginasEnterobius Vermicularis-2011Choi Kio Qa100% (1)
- Conocimiento de La Enfermera en La Atencion A Usuario Con Preeclampsia. Marco ReferencialDocumento10 páginasConocimiento de La Enfermera en La Atencion A Usuario Con Preeclampsia. Marco ReferencialKarlitha Torres EspinoAún no hay calificaciones
- Clase 5 Sap 400 Prevencion, Promocion y Estrategias de PrevencionDocumento38 páginasClase 5 Sap 400 Prevencion, Promocion y Estrategias de PrevencionDorianAún no hay calificaciones
- Dialnet ActualizacionSobreEnfermedadDeMeniere 8950351Documento8 páginasDialnet ActualizacionSobreEnfermedadDeMeniere 8950351Holger BurgosAún no hay calificaciones
- Catalogo VirbacDocumento2 páginasCatalogo VirbacAinoa Bersani RuizAún no hay calificaciones
- Absceso Cerebral para Unsa 2018Documento31 páginasAbsceso Cerebral para Unsa 2018Jonathan Berrio QuintanillaAún no hay calificaciones