También podría gustarte
- Transecto LinealDocumento2 páginasTransecto LinealJose Daniel VasquezAún no hay calificaciones
- La Conducta Manifiesta Al PoderDocumento4 páginasLa Conducta Manifiesta Al PoderLaurita Orozco Álvarez50% (2)
- Proyecto Comunitario. Club de DiabeticosDocumento28 páginasProyecto Comunitario. Club de DiabeticosZharick Narváez100% (3)
- Filogeografia Objetivos Metodos y Ejempl PDFDocumento9 páginasFilogeografia Objetivos Metodos y Ejempl PDFAlejandro Morales GomezAún no hay calificaciones
- Unidad 1 GeneticaDocumento16 páginasUnidad 1 GeneticadiegoAún no hay calificaciones
- Transcripción Del ADNDocumento45 páginasTranscripción Del ADNClauMcClauAún no hay calificaciones
- Protocolo de Extracción de ADNDocumento2 páginasProtocolo de Extracción de ADN40300000% (1)
- Cap 12. Genetica CuantitativaDocumento60 páginasCap 12. Genetica CuantitativaVanessa MelendezAún no hay calificaciones
- Silabo Biologia MolecularDocumento11 páginasSilabo Biologia MolecularRosa Maria Hernàndez RamirezAún no hay calificaciones
- Capitulo 4Documento28 páginasCapitulo 4Samir Gustavo Güiza RomeroAún no hay calificaciones
- Pages From Introduccion A Las Escuelas de Sistematica Y Biogeografia-Scrocchi-1992-2Documento20 páginasPages From Introduccion A Las Escuelas de Sistematica Y Biogeografia-Scrocchi-1992-2Ale MonjeAún no hay calificaciones
- Biología MolecularDocumento41 páginasBiología MolecularRebeca MendozaAún no hay calificaciones
- Biología. La Vida en La Tierra Con Fisiología (9º Edición)Documento9 páginasBiología. La Vida en La Tierra Con Fisiología (9º Edición)Catalina ArayaAún no hay calificaciones
- Genética SchwanDocumento85 páginasGenética SchwanCamila EgasAún no hay calificaciones
- Mecanismo de HeredabilidadDocumento27 páginasMecanismo de HeredabilidadCarlos Xavier Rodriguez100% (2)
- Cruzamientos en Ganado LecheroDocumento6 páginasCruzamientos en Ganado LecheroVenezuelaGanadera.comAún no hay calificaciones
- La Herencia Genética Son Las Características FisiológicasDocumento8 páginasLa Herencia Genética Son Las Características FisiológicasPaola Vianey Gonzalez Navarro100% (1)
- Artículo Revisión BioetanolDocumento11 páginasArtículo Revisión BioetanolCristian ChiiliitoAún no hay calificaciones
- Aporte de La GeneticaDocumento4 páginasAporte de La Geneticajulian smith0% (1)
- Embriologia AnimalDocumento5 páginasEmbriologia Animalarchi90Aún no hay calificaciones
- Manual Laboratorio BQDocumento74 páginasManual Laboratorio BQtony de la rosaAún no hay calificaciones
- CriomicroscopíaDocumento7 páginasCriomicroscopíaedisonpampaAún no hay calificaciones
- MutagénesisDocumento90 páginasMutagénesistalia sartoriAún no hay calificaciones
- Libro Mejoramiento GeneticoDocumento184 páginasLibro Mejoramiento GeneticohenrydjpieroAún no hay calificaciones
- Syllabus Del Curso Virología PDFDocumento11 páginasSyllabus Del Curso Virología PDFEdwin Martinez OrdoñezAún no hay calificaciones
- 57.dominguezetal.2009. Cap - Ephemeroptera MacroinvertebradosBentonicosDocumento45 páginas57.dominguezetal.2009. Cap - Ephemeroptera MacroinvertebradosBentonicosLuis Carlos Villarreal DiazAún no hay calificaciones
- Informe 1 y 2 - Crecimiento y Desarrollo Trabajo en Equipo.Documento14 páginasInforme 1 y 2 - Crecimiento y Desarrollo Trabajo en Equipo.JOSE ANTONI FRAN RAMOS SANCHEZAún no hay calificaciones
- Clasificación de Los Componentes Químicos VegetalesDocumento1 páginaClasificación de Los Componentes Químicos VegetalesVida Vallejiana100% (1)
- TP 4 Genetica de Poblaciones 1Documento32 páginasTP 4 Genetica de Poblaciones 1Fiona MartinezAún no hay calificaciones
- Origen de La Vida, Teorias. Evolucion. CelulaDocumento90 páginasOrigen de La Vida, Teorias. Evolucion. CelulaJossy MoraAún no hay calificaciones
- GenéticaDocumento35 páginasGenéticaANTONIO MEJIAAún no hay calificaciones
- Leyes de MendelDocumento15 páginasLeyes de MendelRosa Amelia Marcelo MairaAún no hay calificaciones
- RAPDDocumento10 páginasRAPDAlejandro MuñozAún no hay calificaciones
- Herencia y Genetica PDFDocumento67 páginasHerencia y Genetica PDFCarla PegueroAún no hay calificaciones
- Técnicas de Cultivo de Microorganismos. Medios y Modelos de Selección, Crecimiento y Mantenimiento.Documento6 páginasTécnicas de Cultivo de Microorganismos. Medios y Modelos de Selección, Crecimiento y Mantenimiento.MarinaAún no hay calificaciones
- Acumulacion Grados Dias - Amanda RomeroDocumento7 páginasAcumulacion Grados Dias - Amanda RomeroFabian TordecillaAún no hay calificaciones
- Manual de Practica de GeneticaDocumento96 páginasManual de Practica de GeneticaDarly Bernabe Rimachi Huaman100% (1)
- Manual BADocumento40 páginasManual BAx4sAún no hay calificaciones
- Conceptos Básicos de Cinética QuímicaDocumento8 páginasConceptos Básicos de Cinética QuímicaNicole Del Solar LagosAún no hay calificaciones
- Principios de La Biologia CelularDocumento22 páginasPrincipios de La Biologia CelularDaniela Espinoza Villegas100% (1)
- Reloj Molecular (Underdog) PDFDocumento47 páginasReloj Molecular (Underdog) PDFhomarmejia1805100% (1)
- CovarianciaDocumento16 páginasCovarianciaMalon Sanchez BenjumeaAún no hay calificaciones
- Marcadores Moleculares y Su Aplicación en PlantasDocumento4 páginasMarcadores Moleculares y Su Aplicación en PlantasDAYANARA ENIHT LIZARAZO ESPINELAún no hay calificaciones
- Ejercicios Del Práctico de PCRDocumento1 páginaEjercicios Del Práctico de PCRManuel Eduardo Peña ZúñigaAún no hay calificaciones
- Introducción A La Bioquímica AgrícolaDocumento9 páginasIntroducción A La Bioquímica AgrícolaMartha SerranoAún no hay calificaciones
- Clave Generic A de Gastropod OsDocumento47 páginasClave Generic A de Gastropod OsKaren PerezAún no hay calificaciones
- Neuroptera ClaseDocumento6 páginasNeuroptera ClaseJerson50% (2)
- Estudio Preliminar de Algunos Aspectos Ambientales y Ecologicos de Peces y Macroinvertebrados en El Rio Tutunendo PDFDocumento10 páginasEstudio Preliminar de Algunos Aspectos Ambientales y Ecologicos de Peces y Macroinvertebrados en El Rio Tutunendo PDFGabo MontesAún no hay calificaciones
- Informe Final de GeneticaDocumento43 páginasInforme Final de GeneticaRosa Mirelly Huaman ChungaAún no hay calificaciones
- Grano de Polen Grupo 9Documento11 páginasGrano de Polen Grupo 9EsmeraldaFlorCaceresPatricioAún no hay calificaciones
- Métodos de Conservación de MicroorganismosDocumento17 páginasMétodos de Conservación de MicroorganismosEmiliano Mojica 1FV1Aún no hay calificaciones
- Ecología Microbiana PDFDocumento51 páginasEcología Microbiana PDFLuciana Sossiré Vásquez Regalado100% (1)
- Preparacion MatrigelDocumento1 páginaPreparacion MatrigelEliud Florentino MolinaAún no hay calificaciones
- Nutricion VegetalDocumento5 páginasNutricion Vegetalwilliam Armando CanacuanAún no hay calificaciones
- Desnaturalización de ProteínasDocumento6 páginasDesnaturalización de ProteínasNANCY FABIOLA ARMERO ALVAREZAún no hay calificaciones
- Proteomica Base de Datos de Proteinas y ProteomasDocumento19 páginasProteomica Base de Datos de Proteinas y ProteomasgonzalofrankAún no hay calificaciones
- Manual para Evaluación BiodiversidadDocumento82 páginasManual para Evaluación BiodiversidadJohn GrahamAún no hay calificaciones
- Practica #10 Sistema Esqueletico de Los PecesDocumento6 páginasPractica #10 Sistema Esqueletico de Los Peces21031b0561Aún no hay calificaciones
- Pensamientos ambientales II: Reflexiones críticas y un tanto irreverentesDe EverandPensamientos ambientales II: Reflexiones críticas y un tanto irreverentesAún no hay calificaciones
- Educación en bioética y alimentos genéticamente modificadosDe EverandEducación en bioética y alimentos genéticamente modificadosAún no hay calificaciones
- Tipos de Anestesia y Fichas FarmacológicasDocumento12 páginasTipos de Anestesia y Fichas FarmacológicasAldo Alva Pérez100% (2)
- Fisiopatología de La Diabetes e IncretinasDocumento6 páginasFisiopatología de La Diabetes e IncretinasmidestinolomejorAún no hay calificaciones
- Presentacion-Músculos de Cortes BovinosDocumento20 páginasPresentacion-Músculos de Cortes BovinosRaul KimAún no hay calificaciones
- Amortiguadores BioquimicaDocumento3 páginasAmortiguadores BioquimicaPaúl FernandoAún no hay calificaciones
- Choice EsplacnoDocumento6 páginasChoice EsplacnoCarlos Somos MediAún no hay calificaciones
- Administracion de Medicamentos Por Via OralDocumento33 páginasAdministracion de Medicamentos Por Via OralYeick Villcas Salazar100% (1)
- Foro 1 AnatomiaDocumento1 páginaForo 1 AnatomiaAngie Carolina BustosAún no hay calificaciones
- Características VocalesDocumento4 páginasCaracterísticas Vocales'Loko Mauri CerpaAún no hay calificaciones
- DG de Histología - Documentos de GoogleDocumento33 páginasDG de Histología - Documentos de GoogleAbril GodiñoAún no hay calificaciones
- 02 TendinomuscularesDocumento58 páginas02 TendinomuscularesPaulina EspinozaAún no hay calificaciones
- Sexologia Definicion EspDocumento30 páginasSexologia Definicion Espivanaeniasaglo100% (2)
- TERMORREGULACIÓNDocumento6 páginasTERMORREGULACIÓNPaulina ZuritaAún no hay calificaciones
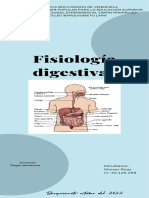
- Infografia Fisiología DigestivaDocumento3 páginasInfografia Fisiología DigestivaMoises RoasAún no hay calificaciones
- TFM M150 PDFDocumento100 páginasTFM M150 PDFPlacido Rojas FrancoAún no hay calificaciones
- Insuficiencia Cardiaca WordDocumento19 páginasInsuficiencia Cardiaca WordJohan Paredes Rodriguez0% (1)
- Hongos LevaduriformesDocumento37 páginasHongos LevaduriformesHilary MendozaAún no hay calificaciones
- Acta de Actividad Fisica AgostoDocumento5 páginasActa de Actividad Fisica AgostoCarlos Magno100% (1)
- Prefijos y SufijosDocumento4 páginasPrefijos y SufijosJG PQ50% (2)
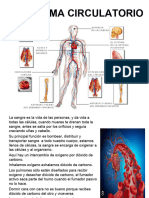
- Sistema Circulatorio 2024Documento24 páginasSistema Circulatorio 2024Sandoval Rodarte DaliaAún no hay calificaciones
- Estrategias Autocuidado Federico Infante LembckeDocumento32 páginasEstrategias Autocuidado Federico Infante LembckeOscar Mendoza LunaAún no hay calificaciones
- Biología 2Documento8 páginasBiología 2Pat SuıcideAún no hay calificaciones
- Child PughDocumento2 páginasChild PughagustinAún no hay calificaciones
- Estudio SomatológicoDocumento2 páginasEstudio Somatológicoareli30100% (2)
- Fundamento Filosofico Del Masaje de Contencion EmocionalDocumento3 páginasFundamento Filosofico Del Masaje de Contencion EmocionalCarolina FauAún no hay calificaciones
- Termoregulacion, Altitud y DopingDocumento11 páginasTermoregulacion, Altitud y DopingDiego CorreaAún no hay calificaciones
- Enzimología Clínica.Documento7 páginasEnzimología Clínica.Lara MayoralAún no hay calificaciones
- Hipertrofia Muscular, Descubre Todas Sus Claves - Mundo EntrenamientoDocumento7 páginasHipertrofia Muscular, Descubre Todas Sus Claves - Mundo EntrenamientoMiguel PadillaAún no hay calificaciones
- El Shock Séptico Es Un Estado de Hipoperfusión Tisular Grave Desencadenado Por La Respuesta Inflamatoria Sistémica de Origen InfecciosoDocumento6 páginasEl Shock Séptico Es Un Estado de Hipoperfusión Tisular Grave Desencadenado Por La Respuesta Inflamatoria Sistémica de Origen Infecciosojaver andres hernandezveraAún no hay calificaciones