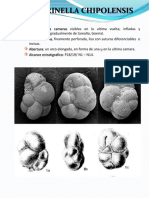
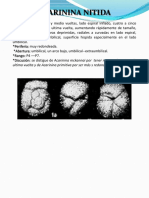
También podría gustarte
- Contrato Compra Venta CocheDocumento1 páginaContrato Compra Venta Cochetomiliet756450% (2)
- Introducción - Perforación IDocumento50 páginasIntroducción - Perforación Iddiazve100% (1)
- BioestratigrafiaDocumento41 páginasBioestratigrafiaPaula Andrea Pacheco100% (3)
- Tipos de FósilesDocumento14 páginasTipos de Fósilesservicioveterinariorottpast995167% (9)
- Miomatosis UterinaDocumento27 páginasMiomatosis UterinaCristian Joshua Hernandez GonzalezAún no hay calificaciones
- Contabilidad Actividad 3 FinalDocumento10 páginasContabilidad Actividad 3 FinalPlayboycorpAún no hay calificaciones
- Ciencia de La Tierra Origen de La TierraDocumento25 páginasCiencia de La Tierra Origen de La Tierraedith arzolaAún no hay calificaciones
- Construcción de Columna EstratigraficaDocumento18 páginasConstrucción de Columna EstratigraficaJose Krlos GüäriinAún no hay calificaciones
- Estratigrafia Js 1Documento100 páginasEstratigrafia Js 1mlcors3244Aún no hay calificaciones
- Cuestionario Examen FinalDocumento5 páginasCuestionario Examen FinalAlex FarfanAún no hay calificaciones
- Geología Aplicada A La HidrogeologíaDocumento29 páginasGeología Aplicada A La HidrogeologíaAllan Ordeñana100% (1)
- MICROPALEONTOLOGIADocumento100 páginasMICROPALEONTOLOGIAJuniors Dueñas LópezAún no hay calificaciones
- Curso Control Geológico-MESOZOICODocumento73 páginasCurso Control Geológico-MESOZOICODaniel C100% (1)
- Tipos de AcuiferosDocumento15 páginasTipos de AcuiferosAlfredo CT100% (1)
- Bioestratigrafia Industria PetroleraDocumento9 páginasBioestratigrafia Industria Petroleraparrakv100% (1)
- Introduccion Al Estudio de Inclusiones Fluidas FA IngenierosDocumento98 páginasIntroduccion Al Estudio de Inclusiones Fluidas FA IngenierosKevin Yunior Mallma PerezAún no hay calificaciones
- 06 - Foraminiferos (2) - 1Documento38 páginas06 - Foraminiferos (2) - 1John LeeAún no hay calificaciones
- Huayacocotla PDFDocumento7 páginasHuayacocotla PDFMauricio ArgüellesAún no hay calificaciones
- Economia CircularDocumento9 páginasEconomia CircularLuis Alirio Angarita OteroAún no hay calificaciones
- La Contaminación Del Aire en MonterreyDocumento7 páginasLa Contaminación Del Aire en MonterreyMarianasancarAún no hay calificaciones
- 1-Informe de RadiolariosDocumento15 páginas1-Informe de RadiolariosMin Nolasco Ibia100% (1)
- Unidades Bioestratigraficas XDDDDDocumento12 páginasUnidades Bioestratigraficas XDDDDRyan Jair Valle QuispeAún no hay calificaciones
- El Compost Como Abono OrganicoDocumento17 páginasEl Compost Como Abono Organicojrarbelo100% (1)
- El Registro de Las RocasDocumento39 páginasEl Registro de Las RocasGael Garcia Martinez100% (1)
- Principios EstratigraficosDocumento15 páginasPrincipios EstratigraficosMarco Alexander PerezAún no hay calificaciones
- Fundamentos de La InvestigaciónDocumento12 páginasFundamentos de La InvestigaciónStephen UbalAún no hay calificaciones
- A4 I.conceptos Sedimentlogicos 23 PDFDocumento53 páginasA4 I.conceptos Sedimentlogicos 23 PDFFrancisco Javier Vazquez Garcia100% (3)
- Fundamento Micropaleontologia 1Documento27 páginasFundamento Micropaleontologia 1Sebas EspínAún no hay calificaciones
- Laboratorio-Introduccion Mapas GeologicosDocumento25 páginasLaboratorio-Introduccion Mapas GeologicosCristian RojasAún no hay calificaciones
- Ostracod OsDocumento2 páginasOstracod OsChris Lp0% (1)
- Sin TítuloDocumento42 páginasSin TítuloWilliamd BautistaAún no hay calificaciones
- 08 Calpionelidos y TintinidosDocumento9 páginas08 Calpionelidos y TintinidosJorge BenitesAún no hay calificaciones
- Tema MicropaleontologíaDocumento196 páginasTema MicropaleontologíaAnderson Vladimir Andrades EscobarAún no hay calificaciones
- 13 Rocas CarbonatadasDocumento28 páginas13 Rocas CarbonatadasJonathanIsraelGalarceMuñozAún no hay calificaciones
- El Jurasico Del SuresteDocumento15 páginasEl Jurasico Del SuresteRafael Diaz RojasAún no hay calificaciones
- Bioestratigrafia SecuencialDocumento116 páginasBioestratigrafia Secuencialanali balvina ccoa yucra100% (1)
- Discontinuidades EstratigraficasDocumento23 páginasDiscontinuidades EstratigraficasNinoshka Deniss Artola MartinezAún no hay calificaciones
- Diccionario de Terminos BotanicosDocumento176 páginasDiccionario de Terminos Botanicosandre rivasAún no hay calificaciones
- Seismic Unix (Ejemplo)Documento23 páginasSeismic Unix (Ejemplo)Federico Gómez González100% (1)
- 1er Respuestas Exam Cuencas U I IIIDocumento10 páginas1er Respuestas Exam Cuencas U I IIICarlos SantanaAún no hay calificaciones
- CLASE MICROPALEONTOLOGIA 2do PARCIAL PDFDocumento65 páginasCLASE MICROPALEONTOLOGIA 2do PARCIAL PDFRene De La OAún no hay calificaciones
- Curso de PaleoambientesDocumento72 páginasCurso de PaleoambientesThanael01100% (1)
- Curso de Geoquimica OrganicaDocumento130 páginasCurso de Geoquimica OrganicaJaor JnrAún no hay calificaciones
- Cap 5 Diatomeas Radiolarios y SilicoflageladosDocumento79 páginasCap 5 Diatomeas Radiolarios y SilicoflageladosBryan Vela Zegarra100% (1)
- Micropaleontologia y PalinologiaDocumento101 páginasMicropaleontologia y PalinologiaJean Pierre Galindo0% (1)
- Calpionelidos y TintinidosDocumento9 páginasCalpionelidos y TintinidosAnahí Carbonell PlatasAún no hay calificaciones
- Maloob-229DES - Programa de Perforación ReducidoDocumento62 páginasMaloob-229DES - Programa de Perforación ReducidoDavid GomezAún no hay calificaciones
- Micro Clase 11 Polen EsporasDocumento79 páginasMicro Clase 11 Polen EsporasDiogo Salvatore100% (1)
- TesisDocumento216 páginasTesisRafael Chi Lopez100% (1)
- Relación Del Entrampamiento de Hidrocarburos y Su ProducciónDocumento16 páginasRelación Del Entrampamiento de Hidrocarburos y Su ProducciónshaireAún no hay calificaciones
- Pricipales Metodos de Analisis Geoquimico PDFDocumento50 páginasPricipales Metodos de Analisis Geoquimico PDFDiego Yana Mendoza88% (16)
- Código de Nomenclatura EstratigráficaDocumento15 páginasCódigo de Nomenclatura EstratigráficaDiego CombitaAún no hay calificaciones
- Estratigrafia SismicaDocumento40 páginasEstratigrafia SismicaAngel SaldañaAún no hay calificaciones
- Definicion y Axiomas de La BiologiaDocumento13 páginasDefinicion y Axiomas de La BiologiaLP JimeAún no hay calificaciones
- Origen Inorgánico Del PetróleoDocumento12 páginasOrigen Inorgánico Del PetróleoJavier RiosAún no hay calificaciones
- DolomiasDocumento13 páginasDolomiasLegabal100% (1)
- Cuenca Tampico MisantlaDocumento46 páginasCuenca Tampico MisantlaOscar Hernandez AlbarranAún no hay calificaciones
- Metodologia Brilha - En.esDocumento16 páginasMetodologia Brilha - En.esCamila MAún no hay calificaciones
- Geoquimica y GeobotanicaDocumento13 páginasGeoquimica y GeobotanicaMelvin GuerraAún no hay calificaciones
- TIF C. CastilloDocumento56 páginasTIF C. CastilloNicolas MirandaAún no hay calificaciones
- Práctica Geol Petrol-Poza Rica 2020-2Documento16 páginasPráctica Geol Petrol-Poza Rica 2020-2DavidAún no hay calificaciones
- El banco de las ánimas: valoración de su biodiversidadDe EverandEl banco de las ánimas: valoración de su biodiversidadAún no hay calificaciones
- Microfósiles Calcáreos GeoDocumento49 páginasMicrofósiles Calcáreos GeoElvs IchevarriaAún no hay calificaciones
- Taxonomía y BacteriasDocumento29 páginasTaxonomía y BacteriasRoosvelt Roberth Ramirez GuillermoAún no hay calificaciones
- Baja Resistividad de AceiteDocumento13 páginasBaja Resistividad de Aceitelalo del angelAún no hay calificaciones
- Globigerina TapuriensisDocumento1 páginaGlobigerina Tapuriensislalo del angelAún no hay calificaciones
- Globigerina TripartitaDocumento1 páginaGlobigerina Tripartitalalo del angelAún no hay calificaciones
- Globigerina OuachitensisDocumento1 páginaGlobigerina Ouachitensislalo del angelAún no hay calificaciones
- Globigerina SelliDocumento1 páginaGlobigerina Sellilalo del angelAún no hay calificaciones
- Globigerina AmpliaperturaDocumento1 páginaGlobigerina Ampliaperturalalo del angelAún no hay calificaciones
- Morozovella SpinulosaDocumento1 páginaMorozovella Spinulosalalo del angelAún no hay calificaciones
- Cribrohantkenina Inflata PDFDocumento1 páginaCribrohantkenina Inflata PDFlalo del angelAún no hay calificaciones
- Globigerina Ciperoensis Angulisuturalis PDFDocumento1 páginaGlobigerina Ciperoensis Angulisuturalis PDFlalo del angelAún no hay calificaciones
- Biocronomarcadores BolliDocumento14 páginasBiocronomarcadores Bollilalo del angelAún no hay calificaciones
- Globigerina Ciperoensis AngustiumbilicataDocumento1 páginaGlobigerina Ciperoensis Angustiumbilicatalalo del angelAún no hay calificaciones
- Globigerina Binaiensis PDFDocumento1 páginaGlobigerina Binaiensis PDFlalo del angelAún no hay calificaciones
- Globigerina Ciperoensis AngulisuturalisDocumento1 páginaGlobigerina Ciperoensis Angulisuturalislalo del angelAún no hay calificaciones
- Globigerina Ciperoensis CiperoensisDocumento1 páginaGlobigerina Ciperoensis Ciperoensislalo del angelAún no hay calificaciones
- Acarinina Bullbrooki PDFDocumento1 páginaAcarinina Bullbrooki PDFlalo del angelAún no hay calificaciones
- Cribrohantkenina InflataDocumento1 páginaCribrohantkenina Inflatalalo del angelAún no hay calificaciones
- Clavatorella BermudeziDocumento1 páginaClavatorella Bermudezilalo del angelAún no hay calificaciones
- Globigerina BinaiensisDocumento1 páginaGlobigerina Binaiensislalo del angelAún no hay calificaciones
- Catapsydrax DissimilisDocumento1 páginaCatapsydrax Dissimilislalo del angelAún no hay calificaciones
- Catapsidrax StainforthiDocumento1 páginaCatapsidrax Stainforthilalo del angelAún no hay calificaciones
- Cassigerinella ChipolensisDocumento1 páginaCassigerinella Chipolensislalo del angelAún no hay calificaciones
- Ejercicio de SobrecargaDocumento3 páginasEjercicio de Sobrecargalalo del angelAún no hay calificaciones
- Acarinina NitidaDocumento1 páginaAcarinina Nitidalalo del angelAún no hay calificaciones
- BiocronomarcadoresDocumento14 páginasBiocronomarcadoreslalo del angelAún no hay calificaciones
- Presentación1 (Autoguardado)Documento1 páginaPresentación1 (Autoguardado)lalo del angelAún no hay calificaciones
- Presentación1 (Autoguardado DigitalDocumento1 páginaPresentación1 (Autoguardado Digitallalo del angelAún no hay calificaciones
- Def1 (Autoguardado DigitalDocumento1 páginaDef1 (Autoguardado Digitallalo del angelAún no hay calificaciones
- Coordinación de VCDSE de Pozos - Luis Carlos PDFDocumento114 páginasCoordinación de VCDSE de Pozos - Luis Carlos PDFlalo del angel50% (2)
- P-SI-005 - V2.docx ProcedimientoDocumento6 páginasP-SI-005 - V2.docx ProcedimientoAmparo Rodriguez GarciaAún no hay calificaciones
- Guía de Ejercicios - Intro A La EconomíaDocumento13 páginasGuía de Ejercicios - Intro A La EconomíaCliff Orlando Bahamondes PobleteAún no hay calificaciones
- Presentación Láser TerapéuticoDocumento18 páginasPresentación Láser TerapéuticoElio Jose Gonzalez BernalAún no hay calificaciones
- Solicitud de Apoyo JudicialDocumento3 páginasSolicitud de Apoyo JudicialMary Gutierrez GilAún no hay calificaciones
- Control Mat - EmpDocumento3 páginasControl Mat - EmpRicardo Ponce De León100% (1)
- Distitron 5119 Elsxq F.T. - EspDocumento1 páginaDistitron 5119 Elsxq F.T. - Espsara.mebautoAún no hay calificaciones
- 126 - Tumores Cutáneos Cervicofaciales Carcinoma Epidermoide y BasocelularDocumento10 páginas126 - Tumores Cutáneos Cervicofaciales Carcinoma Epidermoide y BasocelularCindy FloresAún no hay calificaciones
- Anexo 7 Informe de ITSE PreviaDocumento11 páginasAnexo 7 Informe de ITSE PreviaPamela QhAún no hay calificaciones
- Sig-Pet-Ope-46021-009 Mantenimiento de Chute Alimentacion y DescargaDocumento10 páginasSig-Pet-Ope-46021-009 Mantenimiento de Chute Alimentacion y DescargaIvan Caceres RAún no hay calificaciones
- Práctica #2. Dispositivos de Accionamientos Eléctricos en Automatismos IndustrialesDocumento11 páginasPráctica #2. Dispositivos de Accionamientos Eléctricos en Automatismos IndustrialesDaniel David Mantilla QuinteroAún no hay calificaciones
- Sistemas ModalesDocumento3 páginasSistemas ModalesHumberto Fernando Moncada PeñaAún no hay calificaciones
- Reporte 4 - Movimiento ParabólicoDocumento5 páginasReporte 4 - Movimiento ParabólicoMARCELA CALDERON MORALESAún no hay calificaciones
- Práctica No.12 "Factores Que Afectan La Velocidad de Reacción Enzimática "Documento11 páginasPráctica No.12 "Factores Que Afectan La Velocidad de Reacción Enzimática "Angel Gustavo HernandezAún no hay calificaciones
- Trabajo Final Metodologia IIDocumento31 páginasTrabajo Final Metodologia IIGeraldine AdriaenssensAún no hay calificaciones
- 33 Comunicado UrgenteDocumento6 páginas33 Comunicado UrgenteLucas Olivera PenadilloAún no hay calificaciones
- Organica I (VINO DE MISA)Documento4 páginasOrganica I (VINO DE MISA)Kathia NanamiAún no hay calificaciones
- Calificacion ING. COMERCIAL Verano 3-2021Documento6 páginasCalificacion ING. COMERCIAL Verano 3-2021Rodrigo MeridaAún no hay calificaciones
- Manual Practicas QuimicaDocumento56 páginasManual Practicas QuimicaDiana Ximena García Gámez0% (1)
- Depreciacion Contable Activos FijosDocumento2 páginasDepreciacion Contable Activos FijosElmer RomeroAún no hay calificaciones
- Calculo de La Cantidad de Ladrillos v.1.5 PDFDocumento5 páginasCalculo de La Cantidad de Ladrillos v.1.5 PDFCarlos AyamamaniAún no hay calificaciones
- Comunicar Testimonios de Manera Oral PDFDocumento6 páginasComunicar Testimonios de Manera Oral PDFMaribel de la CruzAún no hay calificaciones
- Bad Bug Book 2nd Edition PDFDocumento310 páginasBad Bug Book 2nd Edition PDFSandra CastilloAún no hay calificaciones
- Líquido Amniótico 2014Documento5 páginasLíquido Amniótico 2014daniloAún no hay calificaciones
- Gestión de Las Cuencas Hidrograficas en EcuadorDocumento20 páginasGestión de Las Cuencas Hidrograficas en EcuadorAndres Macas100% (3)
- CBR InalteradoDocumento4 páginasCBR InalteradoPercy RojasAún no hay calificaciones
- Guia para Los Solicitantes de SubvencionesDocumento27 páginasGuia para Los Solicitantes de SubvencionesVivian Lezama PizzatiAún no hay calificaciones
- Credito Cafetero Estrada y Sandoval Version Publicacion 2014Documento53 páginasCredito Cafetero Estrada y Sandoval Version Publicacion 2014Nasly TatianaAún no hay calificaciones