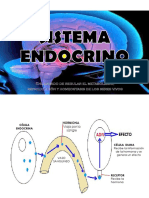
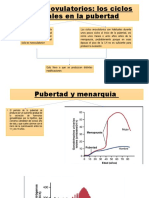
También podría gustarte
- EmbriologiaDocumento62 páginasEmbriologiaendrivall 432Aún no hay calificaciones
- Perfil Hormonal MasculinoDocumento31 páginasPerfil Hormonal MasculinoBrenda DiazAún no hay calificaciones
- RECEPTORESDocumento5 páginasRECEPTORESMaryen Gallardo ovandoAún no hay calificaciones
- Contacto GCDocumento9 páginasContacto GCCarmen Lucía Ferrer PérezAún no hay calificaciones
- Reconocimiento de Las Características Morfológicas y Funcionales Del Aparato Genital FemeninoDocumento15 páginasReconocimiento de Las Características Morfológicas y Funcionales Del Aparato Genital FemeninoGCAún no hay calificaciones
- Endocrinotodo 1Documento79 páginasEndocrinotodo 1Jesus Gabriel CaballeroAún no hay calificaciones
- Ciclo Ovarico, Ovulación, Fecundación, Primera Semana, ImplantaciónDocumento10 páginasCiclo Ovarico, Ovulación, Fecundación, Primera Semana, ImplantaciónAstrid SofíaAún no hay calificaciones
- Gu A BL-11 Ciclo Sexual FemeninoDocumento24 páginasGu A BL-11 Ciclo Sexual FemeninoRafael Videla MontesAún no hay calificaciones
- Prueba Diagnóstica Grado 9 Biología 2021 (Validada)Documento13 páginasPrueba Diagnóstica Grado 9 Biología 2021 (Validada)Juan Pablo Lopez RomeroAún no hay calificaciones
- Ginecología VeterinariaDocumento400 páginasGinecología VeterinariaSan Valentin Agroveterinaria100% (1)
- Dra CaMENOPAUSIA Y HOMEOPATIAlleja Menopausia HomeopatiaDocumento5 páginasDra CaMENOPAUSIA Y HOMEOPATIAlleja Menopausia HomeopatiaVerónica Martínez GonzálezAún no hay calificaciones
- Efecto de Dos Protocolos de Superovulación en Cantidad y Calidad de Embriones Bovinos Producidos in VivoDocumento9 páginasEfecto de Dos Protocolos de Superovulación en Cantidad y Calidad de Embriones Bovinos Producidos in Vivoromel gilberto aguilar valquiAún no hay calificaciones
- Arturo Adolfo Trejos SotoDocumento99 páginasArturo Adolfo Trejos Sotovanessa torresAún no hay calificaciones
- Ginecologia 1er Parcial.Documento176 páginasGinecologia 1er Parcial.Sergio David Vargas ChoqueticllaAún no hay calificaciones
- Taller Sistema EndocrinoDocumento8 páginasTaller Sistema Endocrinomariano padillaAún no hay calificaciones
- Manual de GinecologíaDocumento71 páginasManual de GinecologíaMarsyori FloresAún no hay calificaciones
- Eje Hipotalamo Hipofisis GonadalDocumento8 páginasEje Hipotalamo Hipofisis GonadalIsra BaronaAún no hay calificaciones
- SOLEMNE3Documento48 páginasSOLEMNE3ConiSdafsvthAún no hay calificaciones
- EndocrinoDocumento18 páginasEndocrinoEnrique PuenteAún no hay calificaciones
- ¿Qué Es La Hormona FSH - Valores Normales y Significado Del FSHDocumento14 páginas¿Qué Es La Hormona FSH - Valores Normales y Significado Del FSHAnonymous hgUZlHAún no hay calificaciones
- Espermatogenesis y OvogenesisDocumento15 páginasEspermatogenesis y OvogenesisEva Herrera AhumadaAún no hay calificaciones
- Ciclos AnovulatoriosDocumento7 páginasCiclos Anovulatoriosstefany carreñoAún no hay calificaciones
- Fundamentos Del PCTDocumento4 páginasFundamentos Del PCTLntrainer BodybuilderAún no hay calificaciones
- Preguntas FisiopatologiaDocumento37 páginasPreguntas FisiopatologiaVitor FloreAún no hay calificaciones
- Escalas de TannerDocumento6 páginasEscalas de TannerJean Pierre Cañar ZambranoAún no hay calificaciones
- Deberes Biología 2Documento9 páginasDeberes Biología 2Maria SamaniegoAún no hay calificaciones
- Prueba Coef1 7° Básico Gametos, Regulacion Horrmonal Fem y Masc, Ciclo MenstrualDocumento5 páginasPrueba Coef1 7° Básico Gametos, Regulacion Horrmonal Fem y Masc, Ciclo MenstrualPaola Andrea Valencia Campos80% (5)
- Efectos de Las Hormonas TiroideasDocumento9 páginasEfectos de Las Hormonas TiroideasGabriela LoorAún no hay calificaciones
- Gin-12-16-03 Esterilidad e InfertilidadDocumento9 páginasGin-12-16-03 Esterilidad e InfertilidadOlivera Mollinedo Jerusha NethAún no hay calificaciones
- Anatomia y Fisiologia Del Aparato RepDocumento97 páginasAnatomia y Fisiologia Del Aparato RepCarlos MorilloAún no hay calificaciones