También podría gustarte
- Otras Dinámicas para Aprender La GRATITUDDocumento28 páginasOtras Dinámicas para Aprender La GRATITUDFlor CardenasAún no hay calificaciones
- Infografía Ulcera PépticaDocumento4 páginasInfografía Ulcera PépticaAlex Villanueva100% (1)
- Historia de La GeoestadísticaDocumento9 páginasHistoria de La GeoestadísticaCésar Camacho LojasAún no hay calificaciones
- NORMAs DE SEGURIDAD DE LAS HERRAMIENTAS MANUALEsDocumento5 páginasNORMAs DE SEGURIDAD DE LAS HERRAMIENTAS MANUALEsDanny PrietoAún no hay calificaciones
- Fichas Bonsai BugambillaDocumento11 páginasFichas Bonsai BugambillaJavier AbaloAún no hay calificaciones
- Censo Agropecuario Ine 2013Documento226 páginasCenso Agropecuario Ine 2013Kevin Alexander Chavez ClarosAún no hay calificaciones
- Zanahoria y Uvilla: Los Aderezos de La AbuelaDocumento7 páginasZanahoria y Uvilla: Los Aderezos de La AbuelaCecilia VillacisAún no hay calificaciones
- San Juan Bosco - La Víbora (La Habana) Salesianos de CubaDocumento3 páginasSan Juan Bosco - La Víbora (La Habana) Salesianos de CubaOsvaldo VieraAún no hay calificaciones
- Sierra Maestra 28-03-2020Documento8 páginasSierra Maestra 28-03-2020GabrielAún no hay calificaciones
- Arquitectura Basica de Un Sistema Minimo de ComutadoraDocumento23 páginasArquitectura Basica de Un Sistema Minimo de ComutadoralinchmxAún no hay calificaciones
- El Nabo GiganteDocumento2 páginasEl Nabo GigantejavierAún no hay calificaciones
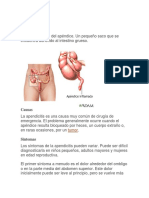
- ApendicitisDocumento7 páginasApendicitisRuli TosmagAún no hay calificaciones
- Toda La Historia Del PerúDocumento55 páginasToda La Historia Del PerúluliAún no hay calificaciones
- PIP RaindropTechniqueDocumento1 páginaPIP RaindropTechniqueMucia RodriguezAún no hay calificaciones
- Terapia PeriodontalDocumento25 páginasTerapia PeriodontalRobert AucancelaAún no hay calificaciones
- 5 Guia Lab. Tratamientos Termicos-2019-1Documento8 páginas5 Guia Lab. Tratamientos Termicos-2019-1Kayla MendozaAún no hay calificaciones
- Informe Empalmes ElectricosDocumento8 páginasInforme Empalmes ElectricosSilvita CarolinaAún no hay calificaciones
- El Origen Del CáncerDocumento8 páginasEl Origen Del CáncerYuliana SotoAún no hay calificaciones
- Ortografia Acentual y Literal LENGUAJE Literatura RafaelBorges 1°MEDIO 166COPIASDocumento3 páginasOrtografia Acentual y Literal LENGUAJE Literatura RafaelBorges 1°MEDIO 166COPIASRafael BorgesAún no hay calificaciones
- 50 Actividades para Trabajar y Estimular La Motricidad InfantilDocumento3 páginas50 Actividades para Trabajar y Estimular La Motricidad InfantilRoxanna MuñozAún no hay calificaciones
- Estudio Socioeconómico de Un ProyectoDocumento56 páginasEstudio Socioeconómico de Un ProyectoRubenSilvaAún no hay calificaciones
- 33 38 EsDocumento35 páginas33 38 Es12joseeligioAún no hay calificaciones
- CatalogoDocumento20 páginasCatalogoMeli CastañoAún no hay calificaciones
- Tabla de Calorías PDFDocumento6 páginasTabla de Calorías PDFMarian AmoresAún no hay calificaciones
- INFORME DE LABORATORIO#7 S2P1 (FACTORES DE INERCIA A LA ROTACIÓN Il)Documento6 páginasINFORME DE LABORATORIO#7 S2P1 (FACTORES DE INERCIA A LA ROTACIÓN Il)Jofre CastilloAún no hay calificaciones
- Gen Eng Spe 002Documento6 páginasGen Eng Spe 002Jose C. MelendezAún no hay calificaciones
- AmeDocumento104 páginasAmesergio honores0% (1)
- Problemas mn1-2Documento4 páginasProblemas mn1-2Junior C. Hurtado0% (1)
- Cotizacion Corregido Brayan FinalDocumento4 páginasCotizacion Corregido Brayan FinalBrayan Motta RodríguezAún no hay calificaciones
- Determinación de ConjuntosDocumento3 páginasDeterminación de ConjuntosAlison Nicole Acosta Alejo0% (1)