También podría gustarte
- Nom 087 ResumenDocumento2 páginasNom 087 ResumenRox SalazarAún no hay calificaciones
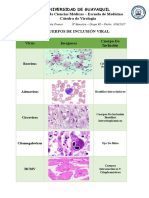
- Universidad de Guayaquil Facultad de Ciencias Médicas - Escuela de Medicina Cátedra de VirologíaDocumento4 páginasUniversidad de Guayaquil Facultad de Ciencias Médicas - Escuela de Medicina Cátedra de VirologíaGenesis GarciaAún no hay calificaciones
- Trabajo MicroorganismosDocumento15 páginasTrabajo MicroorganismosEstibaliz Mechoso MartinezAún no hay calificaciones
- Bases Biologicas de La ConductaDocumento33 páginasBases Biologicas de La ConductaOscar DaríoAún no hay calificaciones
- Marco TeoricoDocumento9 páginasMarco TeoricoThalia HuapayaAún no hay calificaciones
- Glosario BiologiaDocumento53 páginasGlosario BiologiaYohana Andrea Rojas RiañoAún no hay calificaciones
- Glándulas Accesorias Del Sistema DigestivoDocumento21 páginasGlándulas Accesorias Del Sistema Digestivokarly echeverryAún no hay calificaciones
- Librodecienciasnaturales7mogrado 150525140740 Lva1 App6892Documento244 páginasLibrodecienciasnaturales7mogrado 150525140740 Lva1 App6892Cuarto Básico Nery Dos100% (1)
- Diapositivas Flora y Fauna WilderDocumento8 páginasDiapositivas Flora y Fauna WilderWilder Ramírez RamosAún no hay calificaciones
- Guías Resueltas Embrio y GenéticaDocumento103 páginasGuías Resueltas Embrio y GenéticaDaii RamirezAún no hay calificaciones
- Sistema MuscularDocumento14 páginasSistema MuscularDANIEL ALEJANDROAún no hay calificaciones
- Antologia Biologia y Comportamiento PDFDocumento187 páginasAntologia Biologia y Comportamiento PDFRebe O DesAún no hay calificaciones
- Antibióticos Alimentos y AmbienteDocumento4 páginasAntibióticos Alimentos y AmbienteCacho BacacelaAún no hay calificaciones
- Biodiversidad en El PerúDocumento4 páginasBiodiversidad en El PerúmichelAún no hay calificaciones
- Cuestionario HistoDocumento3 páginasCuestionario HistoAlejandro TrenemanAún no hay calificaciones
- Interacción Entre Las Células Gliales y Neuronales y Su Papel en La Muerte y Sobrevivencia NeuronalDocumento9 páginasInteracción Entre Las Células Gliales y Neuronales y Su Papel en La Muerte y Sobrevivencia NeuronalCamilla CristinaAún no hay calificaciones
- Actividad de FenotipoDocumento3 páginasActividad de FenotipoandreaprofeAún no hay calificaciones
- 02 - Mapa Mental - Francisco YánezDocumento1 página02 - Mapa Mental - Francisco YánezLuis Carlos Yánez MarroAún no hay calificaciones
- VIRUSDocumento17 páginasVIRUSL C100% (1)
- Examen Biologia Zonal 2019.correctoDocumento14 páginasExamen Biologia Zonal 2019.correctoEdrey EspinozaAún no hay calificaciones
- Domesticacion - EtologiaDocumento38 páginasDomesticacion - EtologiaFelipe WolfAún no hay calificaciones
- Clase Virtual No. 2. Medios de Cultivo y Obtención de Cultivos Puros PDFDocumento25 páginasClase Virtual No. 2. Medios de Cultivo y Obtención de Cultivos Puros PDFjuan sebastian campo pachecoAún no hay calificaciones
- Linea Del Tiempo Historia de La GeneticaDocumento1 páginaLinea Del Tiempo Historia de La GeneticaLUISA FERNANDA MONCADA GOMEZ100% (6)
- Estructura Externa de La BacteriaDocumento10 páginasEstructura Externa de La BacteriaHoss SeeeAún no hay calificaciones
- Biología CriminalDocumento2 páginasBiología CriminalKevin Mendez100% (1)
- Cancer de OvarioDocumento78 páginasCancer de OvarioALVARO MEDINA100% (1)
- Marco Teorico - Practica 1Documento4 páginasMarco Teorico - Practica 1Carlos Llicán CenturiónAún no hay calificaciones
- CIRROSISDocumento10 páginasCIRROSISedwinmapaAún no hay calificaciones
- Portafolio 3Documento1 páginaPortafolio 3lanchAún no hay calificaciones
- Modificación de Sanger en Capilar.Documento12 páginasModificación de Sanger en Capilar.Luis Ángel CruzAún no hay calificaciones