También podría gustarte
- Cuidados y Rol Del Tens en Un Paciente Con Accidente Cerebro VascularDocumento1 páginaCuidados y Rol Del Tens en Un Paciente Con Accidente Cerebro VascularManuel Vidal Lagos40% (5)
- Plan de Intervención Paciente Postrado Grave y TotalDocumento3 páginasPlan de Intervención Paciente Postrado Grave y TotalManuel Vidal Lagos0% (1)
- TubosDocumento1 páginaTubosManuel Vidal Lagos50% (2)
- MahadutaDocumento17 páginasMahadutaManuel Vidal LagosAún no hay calificaciones
- Circo de Las MariposasDocumento8 páginasCirco de Las MariposasVctr MaderaAún no hay calificaciones
- Funcion de La Nutricion en Los AnimalesDocumento10 páginasFuncion de La Nutricion en Los AnimalesMaico Sanchez la CruzAún no hay calificaciones
- Silabo Fisiologia Vegetal 2013-IIDocumento4 páginasSilabo Fisiologia Vegetal 2013-IIMario Junior Zegarra VásquezAún no hay calificaciones
- Estandarización Del Proceso de Fermentación de Leche Entera Ultrapasteurizada Con Gránulos de KefirDocumento168 páginasEstandarización Del Proceso de Fermentación de Leche Entera Ultrapasteurizada Con Gránulos de KefirBrandito MPAún no hay calificaciones
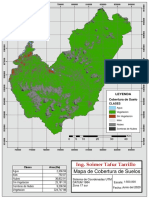
- Mapa de Cobertura de Suelo - UtcubambaDocumento1 páginaMapa de Cobertura de Suelo - UtcubambaSoimer TafurAún no hay calificaciones
- Identificacion Del Funcionamiento de La Celula XDocumento21 páginasIdentificacion Del Funcionamiento de La Celula XEmir CastillaAún no hay calificaciones
- Fibras de Origen VegetalDocumento2 páginasFibras de Origen VegetalDante Lopez Murga100% (1)
- The Evolution of Insect Body Coloration Under Changing Climates - En.esDocumento8 páginasThe Evolution of Insect Body Coloration Under Changing Climates - En.esCristian MonterreyAún no hay calificaciones
- Factores Biologicos de La Criminalidad DiapoDocumento28 páginasFactores Biologicos de La Criminalidad DiapoAstrid EstefaniaAún no hay calificaciones
- Nuevos Agentes RemineralizadoresDocumento59 páginasNuevos Agentes RemineralizadoresKathya AlessandrhaAún no hay calificaciones
- Estudio de Familia UnamDocumento28 páginasEstudio de Familia UnamMyck Stark Ambriz100% (2)
- Guia de Ciencias 1 Semana 29 04 Al 08 OctubreDocumento2 páginasGuia de Ciencias 1 Semana 29 04 Al 08 Octubresandra moraguesAún no hay calificaciones
- Un Viaje Hacia La VerdadDocumento2 páginasUn Viaje Hacia La VerdadAngela Correa100% (1)
- Las PGPRDocumento1 páginaLas PGPRlisett pereyraAún no hay calificaciones
- Embriogénesis Del Sistema NerviosoDocumento12 páginasEmbriogénesis Del Sistema Nerviosoelgrupodetesis7035100% (6)
- Contenido de Informe Mensual de ObraDocumento12 páginasContenido de Informe Mensual de ObraJorge RenattoAún no hay calificaciones
- Manual Dendroctonus SinaloaDocumento22 páginasManual Dendroctonus SinaloaRobert CrumbAún no hay calificaciones
- Afecciones Benignas Del Cuello UterinoDocumento4 páginasAfecciones Benignas Del Cuello UterinoRaimarys GomezAún no hay calificaciones
- Proyectos I+D SENACYT (Hasta Diciembre, 2019)Documento60 páginasProyectos I+D SENACYT (Hasta Diciembre, 2019)Santiago Del CastilloAún no hay calificaciones
- Pubalgia PDFDocumento6 páginasPubalgia PDFRobertto SanchezAún no hay calificaciones
- A Los Últimos 570 Millones de AñosDocumento7 páginasA Los Últimos 570 Millones de AñosManuel RodriguezAún no hay calificaciones
- ELABORACION DE QUESO FRESCO Ing. FocDocumento11 páginasELABORACION DE QUESO FRESCO Ing. FocMp PatriciaAún no hay calificaciones
- 50 Anos de La Doble Helice La Molecula Mas Bella Del MundoDocumento7 páginas50 Anos de La Doble Helice La Molecula Mas Bella Del Mundoapi-264033309Aún no hay calificaciones
- Información Del SidaDocumento4 páginasInformación Del SidaYorman SaulAún no hay calificaciones
- Alumbramiento FisiologicaDocumento31 páginasAlumbramiento Fisiologicaleomar polloAún no hay calificaciones
- Morfología de La Caña de Azúcar en La Preparación Profunda Del Suelo en CanterosDocumento8 páginasMorfología de La Caña de Azúcar en La Preparación Profunda Del Suelo en CanterosCarlosAún no hay calificaciones
- EspagiriaDocumento853 páginasEspagiriaturiel50% (2)
- Zingiberaceae 123Documento6 páginasZingiberaceae 123KellyJohanaAún no hay calificaciones
- Evaluación Unidad 0 Marzo Lenguaje 6°Documento7 páginasEvaluación Unidad 0 Marzo Lenguaje 6°maria piaAún no hay calificaciones
- Actividad EnzimaticaDocumento10 páginasActividad EnzimaticaMelissaChavaAún no hay calificaciones