También podría gustarte
- La Química de Los SentidosDocumento2 páginasLa Química de Los SentidosHector RojasAún no hay calificaciones
- Slide Final de MicroDocumento65 páginasSlide Final de MicroNara FlosanAún no hay calificaciones
- Músculo CardíacoDocumento15 páginasMúsculo CardíacoNara FlosanAún no hay calificaciones
- CONTENIDO FISIOLOGIA 1 (Potencial de Acción Muscular)Documento7 páginasCONTENIDO FISIOLOGIA 1 (Potencial de Acción Muscular)Nara FlosanAún no hay calificaciones
- Juanivan92 Editora de Seccin 6. LM Apgar Familiar VeraDocumento3 páginasJuanivan92 Editora de Seccin 6. LM Apgar Familiar VeraNara FlosanAún no hay calificaciones
- La Música BarrocaDocumento10 páginasLa Música BarrocaAlejandro RodríguezAún no hay calificaciones
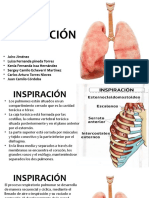
- InspiracionDocumento17 páginasInspiracionKaren Benita Mena PastranaAún no hay calificaciones
- Tratamiento Labio Leporino y Paladar HendidoDocumento5 páginasTratamiento Labio Leporino y Paladar Hendidoximenna nicold vargas arriagaAún no hay calificaciones
- Tejido EpitelialDocumento6 páginasTejido EpitelialStefania FusariAún no hay calificaciones
- Banco Historico-Cirugia General - Nivel Intermedio - Sin ClavesDocumento6 páginasBanco Historico-Cirugia General - Nivel Intermedio - Sin Clavescamportugal27Aún no hay calificaciones
- Capitulo 17 Examen Del AbdomenDocumento38 páginasCapitulo 17 Examen Del AbdomenLuisdeLeonAún no hay calificaciones
- Morfofisiología Bucal Se01Documento7 páginasMorfofisiología Bucal Se01Daniela León100% (2)
- Objetivos de La Asignatura HistologíaDocumento2 páginasObjetivos de La Asignatura Histologíaedgar dominguezAún no hay calificaciones
- Generalidades TACDocumento22 páginasGeneralidades TACCoral LunaAún no hay calificaciones
- Ana - Um4 2022Documento21 páginasAna - Um4 2022X legionAún no hay calificaciones
- MANEJO DE LA VIA AREA EN EL ENTORNO TACTICO - 4 VersiónDocumento25 páginasMANEJO DE LA VIA AREA EN EL ENTORNO TACTICO - 4 VersiónAlvaro PemartinAún no hay calificaciones
- Sistema Nervioso para Quinto de PrimariaDocumento6 páginasSistema Nervioso para Quinto de PrimariaMartha CarvajalAún no hay calificaciones
- Propulsión y Mezcla de Los Alimentos en El Aparato DigestivoDocumento49 páginasPropulsión y Mezcla de Los Alimentos en El Aparato DigestivoOscar1CarreonAún no hay calificaciones
- Regiones Anatómicas Del Cuerpo HumanoDocumento4 páginasRegiones Anatómicas Del Cuerpo HumanoDanek Sirio67% (3)
- El Cuerpo Humano 2021 257 - argMSCDocumento24 páginasEl Cuerpo Humano 2021 257 - argMSCJohana Iafrancisco100% (1)
- Programación Didáctica Anatomia I-Pac-2020 (Sylabo) Modalidad VirtualDocumento54 páginasProgramación Didáctica Anatomia I-Pac-2020 (Sylabo) Modalidad VirtualOscar Waldemar Zschocher CasildoAún no hay calificaciones
- 2 Biología SextoDocumento6 páginas2 Biología Sextoparkour parkourAún no hay calificaciones
- Semana 1 Consolidación 1: para Aprender A Estudiar Que Aparece en Tu CDDocumento22 páginasSemana 1 Consolidación 1: para Aprender A Estudiar Que Aparece en Tu CDyosman lópezAún no hay calificaciones
- 3-2 - Diagnostico Diferencial en Pelvis y CaderaDocumento26 páginas3-2 - Diagnostico Diferencial en Pelvis y CaderaRichard Armijo GarridoAún no hay calificaciones
- RúbricasDocumento8 páginasRúbricasDianelly Colli EkAún no hay calificaciones
- Sistema VenosoDocumento3 páginasSistema VenosoANDRADE CHIRINOS ANGELAAún no hay calificaciones
- Yerly Trabajo Caso Clinico Aorta Abdominal UnadDocumento14 páginasYerly Trabajo Caso Clinico Aorta Abdominal UnadninyAún no hay calificaciones
- AyudinDocumento13 páginasAyudinYocelin FernándezAún no hay calificaciones
- Psicología FisiológicaDocumento7 páginasPsicología FisiológicaAnali Liliana Bayona AcuñaAún no hay calificaciones
- Presentacion de NeuroanatomiaDocumento22 páginasPresentacion de NeuroanatomiaIsis Palacios MoranAún no hay calificaciones
- AnalogíaDocumento6 páginasAnalogíajuanAún no hay calificaciones
- Tejido Conectivo General Laxo y DensoDocumento2 páginasTejido Conectivo General Laxo y DensoMariana RodriguezAún no hay calificaciones
- Irrigación MSDocumento14 páginasIrrigación MSJessica Contreras SandovalAún no hay calificaciones
- Modulo - Anatomia - CEPRE I - 2021 UNDACDocumento32 páginasModulo - Anatomia - CEPRE I - 2021 UNDACGiuliana AlvaradoAún no hay calificaciones