También podría gustarte
- Ácidos Grasos 2Documento5 páginasÁcidos Grasos 2cesarAún no hay calificaciones
- Ejercicio 1Documento2 páginasEjercicio 1cesarAún no hay calificaciones
- Química Orgánica 1Documento5 páginasQuímica Orgánica 1cesarAún no hay calificaciones
- Ácidos Grasos 2Documento5 páginasÁcidos Grasos 2cesarAún no hay calificaciones
- Agil 15Documento16 páginasAgil 15cesarAún no hay calificaciones
- Clasificación Según Los Grupos FuncionalesDocumento10 páginasClasificación Según Los Grupos FuncionalescesarAún no hay calificaciones
- Agil 12Documento4 páginasAgil 12cesarAún no hay calificaciones
- Agile 1Documento6 páginasAgile 1cesarAún no hay calificaciones
- Célula 7Documento5 páginasCélula 7cesarAún no hay calificaciones
- Agil 10Documento3 páginasAgil 10cesarAún no hay calificaciones
- Agil 11Documento6 páginasAgil 11cesarAún no hay calificaciones
- Agile 2Documento11 páginasAgile 2cesarAún no hay calificaciones
- Agil 13Documento9 páginasAgil 13cesarAún no hay calificaciones
- Agil 14Documento3 páginasAgil 14cesarAún no hay calificaciones
- Agile 4Documento4 páginasAgile 4cesarAún no hay calificaciones
- Agile 9Documento2 páginasAgile 9cesarAún no hay calificaciones
- Agile 7Documento7 páginasAgile 7cesarAún no hay calificaciones
- La Medicina-Que EstudiarDocumento2 páginasLa Medicina-Que EstudiarcesarAún no hay calificaciones
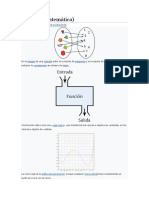
- FunciónDocumento27 páginasFuncióncesarAún no hay calificaciones
- Célula 4Documento2 páginasCélula 4cesarAún no hay calificaciones
- Célula 7Documento5 páginasCélula 7cesarAún no hay calificaciones
- Biología InformacionDocumento10 páginasBiología InformacioncesarAún no hay calificaciones
- Célula: para Otros Usos de Este Término, VéaseDocumento11 páginasCélula: para Otros Usos de Este Término, VéasecesarAún no hay calificaciones
- Cas 077 2020Documento3 páginasCas 077 2020Kathe RpzAún no hay calificaciones
- Gas Natural OsinergDocumento3 páginasGas Natural OsinergcesarAún no hay calificaciones
- Teoría CelularDocumento3 páginasTeoría CelularYesica BlancoAún no hay calificaciones
- Célula 3Documento5 páginasCélula 3cesarAún no hay calificaciones
- CélulaDocumento5 páginasCélulaYesica BlancoAún no hay calificaciones
- Osinerg SupervisionDocumento6 páginasOsinerg SupervisioncesarAún no hay calificaciones
- Guias de Clase Practicas y Seminarios 2Documento24 páginasGuias de Clase Practicas y Seminarios 2Leandro Morales NaranjoAún no hay calificaciones
- Núcleo InterfasicoDocumento14 páginasNúcleo InterfasicoPaolaAún no hay calificaciones
- CICLO CELULAR. Mitosis y MeiosisDocumento21 páginasCICLO CELULAR. Mitosis y MeiosisVengadores AvengersAún no hay calificaciones
- Meiosis 1 y 2Documento2 páginasMeiosis 1 y 2MATIAS NICOLAS ALVAREZ CASTILLOAún no hay calificaciones
- Resumen Biologia (Cooper + Seminarios)Documento100 páginasResumen Biologia (Cooper + Seminarios)La Pecera CopiasAún no hay calificaciones
- Guia 3 Procesos Celulares PDFDocumento26 páginasGuia 3 Procesos Celulares PDFcamila garciaAún no hay calificaciones
- Biologia - George H FriedDocumento10 páginasBiologia - George H FriedRODRÍGUEZ ALEJANDROAún no hay calificaciones
- Preguntas Sobre Meiosis y GamatogenesisDocumento4 páginasPreguntas Sobre Meiosis y GamatogenesisJenny Castro JaraAún no hay calificaciones
- Prometafase, Metafase, Telofase...Documento5 páginasPrometafase, Metafase, Telofase...MICHELLE PATRICIA FAJARDO SIGUENCIAAún no hay calificaciones
- Tarea 2 - Ciclo CelularDocumento3 páginasTarea 2 - Ciclo CelularVanessa PaezAún no hay calificaciones
- Semejanzas y Diferencias Entre Mitosis - MeiosisDocumento10 páginasSemejanzas y Diferencias Entre Mitosis - MeiosisEider SantosAún no hay calificaciones
- Semana-11 Proc BiolDocumento32 páginasSemana-11 Proc BiolLuis OrtizAún no hay calificaciones
- LAB BIO Ejercicios Mitosis y MeiosisDocumento3 páginasLAB BIO Ejercicios Mitosis y MeiosisLian TapiaAún no hay calificaciones
- Fases de La MitosisDocumento2 páginasFases de La MitosisRaffa Pérez100% (1)
- Cuestionario Mitosis y MeiosisDocumento6 páginasCuestionario Mitosis y MeiosisErick Martin Villalobos GomezAún no hay calificaciones
- Biología Celular Ciclo Celular 13 16Documento42 páginasBiología Celular Ciclo Celular 13 16Mateo CCAún no hay calificaciones
- 50 PreguntasDocumento21 páginas50 PreguntasJeni Campos RamosAún no hay calificaciones
- MeiosisDocumento31 páginasMeiosissaaAún no hay calificaciones
- Division Celular MeiosisDocumento6 páginasDivision Celular MeiosisJainely BarbosaAún no hay calificaciones
- Gancino A PRÁCTICA #14 MITOSISDocumento9 páginasGancino A PRÁCTICA #14 MITOSISAndy Gancino0% (1)
- Organizador Meiosis y GametogenésisDocumento1 páginaOrganizador Meiosis y GametogenésisKarla Fabiana CORDOVA OJEDAAún no hay calificaciones
- Guía N°2 - Ciclo Celular - MITOSIS - II-medioDocumento9 páginasGuía N°2 - Ciclo Celular - MITOSIS - II-medioregulus blackAún no hay calificaciones
- II-17 Guía 54 2da ParteDocumento23 páginasII-17 Guía 54 2da ParteAnonymous vxSpL3XBn100% (1)
- Evaluación Biología 8Documento2 páginasEvaluación Biología 8Giancarlo Hernandez PapaleoAún no hay calificaciones
- Ciclo Celular 2Documento46 páginasCiclo Celular 2Karen AvilésAún no hay calificaciones
- La Mitosis y MeiosisDocumento9 páginasLa Mitosis y MeiosisKaren JazmínAún no hay calificaciones
- Ciclo CelularDocumento49 páginasCiclo CelularVirginia BoltenAún no hay calificaciones
- Guía de Trabajo 6. Mitosis - Meiosis 2021-1Documento8 páginasGuía de Trabajo 6. Mitosis - Meiosis 2021-1Luisa MuñozAún no hay calificaciones
- Division CelularDocumento38 páginasDivision CelularAlex QuicenoAún no hay calificaciones
- Trabajo de Biologia ValeDocumento6 páginasTrabajo de Biologia ValeDaniel LopezAún no hay calificaciones