También podría gustarte
- Guia Autoaprendizaje Estudiante 8vo Grado Ciencia f2 s3Documento5 páginasGuia Autoaprendizaje Estudiante 8vo Grado Ciencia f2 s3Missael MartínezAún no hay calificaciones
- Regulación de La Expresión Génica en ProcariontesDocumento32 páginasRegulación de La Expresión Génica en ProcariontesBenjamin Padilla ParodiAún no hay calificaciones
- 20-Estructura Fina Del Gen-Concepto de GenDocumento6 páginas20-Estructura Fina Del Gen-Concepto de GenAlmudena MeiAún no hay calificaciones
- 17-Ligamiento y Recombinación en EucariontesDocumento25 páginas17-Ligamiento y Recombinación en EucariontesAlmudena MeiAún no hay calificaciones
- 17b-Ligamiento y Recombinación en HongosDocumento9 páginas17b-Ligamiento y Recombinación en HongosAlmudena MeiAún no hay calificaciones
- 17b-Ligamiento y Recombinación en HongosDocumento9 páginas17b-Ligamiento y Recombinación en HongosAlmudena MeiAún no hay calificaciones
- 15-Base Genética de La Variación ContinuaDocumento22 páginas15-Base Genética de La Variación ContinuaAlmudena MeiAún no hay calificaciones
- 13-El Polihíbrido. Análisis Estadístico Aplicado La MendelismoDocumento12 páginas13-El Polihíbrido. Análisis Estadístico Aplicado La MendelismoAlmudena MeiAún no hay calificaciones
- BIOQUIMICA Examen Final PDFDocumento217 páginasBIOQUIMICA Examen Final PDFTyAún no hay calificaciones
- Las Moleculas de Adhesion y La Respuesta Inmune PDFDocumento8 páginasLas Moleculas de Adhesion y La Respuesta Inmune PDFAlexa BuenoAún no hay calificaciones
- RT PCRDocumento5 páginasRT PCRAnatomia 2018Aún no hay calificaciones
- Inhibicion CompetitivaDocumento9 páginasInhibicion Competitivakarla torrezAún no hay calificaciones
- EndocrinologíaDocumento35 páginasEndocrinologíaPablo NovoaAún no hay calificaciones
- La Logica MolecularDocumento8 páginasLa Logica MolecularRicardo Jesus Crespo MezaAún no hay calificaciones
- Metabolismo de Los FármacosDocumento17 páginasMetabolismo de Los FármacosVanessa L. Hernández Araujo100% (1)
- Tema - 4 - Estructura Tridimensional de ProteínasDocumento53 páginasTema - 4 - Estructura Tridimensional de ProteínasAitor PastorAún no hay calificaciones
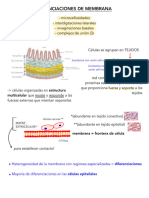
- Diferenciaciones de La Membrana PlasmáticaDocumento5 páginasDiferenciaciones de La Membrana PlasmáticaLou ClementAún no hay calificaciones
- Practica 5 Pruebas de Amilasa y ProteasaDocumento5 páginasPractica 5 Pruebas de Amilasa y ProteasaAngie Borbor100% (1)
- LevadurasDocumento6 páginasLevadurasAngie Santillán ValquiAún no hay calificaciones
- Cancer de PulmonDocumento160 páginasCancer de Pulmontrazavia81100% (1)
- ParacetamolDocumento20 páginasParacetamolVergaraCortezJaredAún no hay calificaciones
- CICLO DE KREBS Power PointDocumento11 páginasCICLO DE KREBS Power Pointjuan pablo100% (1)
- EDIT Curva Estándar. 53E3 - Eunice VillegasDocumento7 páginasEDIT Curva Estándar. 53E3 - Eunice VillegasMarco antonio Sanchez medinaAún no hay calificaciones
- SDHDocumento7 páginasSDHRaul BasilioAún no hay calificaciones
- Banco de Preguntas - ProteínasDocumento2 páginasBanco de Preguntas - ProteínasConsuelo Tamay Cieza100% (2)
- Biologia - 614 - Tarea - 1 - Jorge Enrique Farfan LopezDocumento10 páginasBiologia - 614 - Tarea - 1 - Jorge Enrique Farfan LopezJorge Enrique Farfan LopezAún no hay calificaciones
- Pec 2022-23Documento7 páginasPec 2022-23Rina Horton100% (1)
- 2d Determinacion de ProteinasDocumento29 páginas2d Determinacion de ProteinasAleja Gutiérrez CossioAún no hay calificaciones
- Teorías VirologíaDocumento10 páginasTeorías VirologíaLizeth Aragon GomezAún no hay calificaciones
- Final Bioquimica FMED UBA 3-5-19 TEMA UDocumento3 páginasFinal Bioquimica FMED UBA 3-5-19 TEMA UCande AmiconeAún no hay calificaciones
- Carbohidratos Mapa ConceptualDocumento1 páginaCarbohidratos Mapa ConceptualJosmar TorresAún no hay calificaciones
- ToxicologíaDocumento8 páginasToxicologíajuan jose jasso huamanAún no hay calificaciones
- Cultivo Celular PDFDocumento12 páginasCultivo Celular PDFAdrian VargasAún no hay calificaciones
- Informe Roya Del CafeDocumento19 páginasInforme Roya Del CafeOscar GaloAún no hay calificaciones
- El Zinc en La Salud HumanaDocumento5 páginasEl Zinc en La Salud HumanaSILVIA ALVAREZ RAMIREZAún no hay calificaciones
- Bioquimica LibroDocumento10 páginasBioquimica LibroAngely Dayana Feliciano RamosAún no hay calificaciones
- Biologia Nov 3 SoniaDocumento5 páginasBiologia Nov 3 Soniaysrrael privAún no hay calificaciones