ervation én
2004; 29;
yf erty ‘
Glin Oncol Biologia celular y molecular de la maduracion
ons forthe ovocitaria en mamiferos
= Silvia E, Racedo!, Daniel F Salamone! y Vanesa Y. Rawe?
Society for ‘Laboratorio de Botecaloga Avimal, Focultod de Agronomia, Univerided de Buenos Aves
a ear SLAbiNes, EGR
for ancer
,
——
induced
iol Reprod
in patients BREVES ASPECTOS MOLECULARES DE LA
*armacol FOLICULOGENESIS Y OVOGENESIS.
En especies monovulares,la ovogénesis se podria culogénesis. Durante ese perfodo y demanera coond-
1 oocyte dividir en tres periodos: proliferacién, crecimiento y nada con la ovogénesis, el foliculo primordial pasa a
maduraciOn. Durante la proiferacién tienen lugardi- ser primario, luego secundario y finalmente terciario
prynitee visiones mit6ticas de las ovogonias. En ella sedeter- antral (Rodriguez y Farin, 2004) (Figura 1).
re ‘ina el miimero total decélulas germinales delahem- —_Elejeendoctino-hipotélamo-hipéfiso-gonadal regula
phoma Dra y como consecuencia os ovocitos que serén utill- jos eventos que llevan ala produccicn de pametas enel
ts zados durante la vida reproductiva. El segundo pe- macho y en la hembra. Los foliculos ovisicos son esti=
s6-206 odo corresponde al creimiento del ovocto. En esta. mulados por gonadotrofina (L!1y FSD sntetizacasy
etapa no sélo la eéhula germinal adquiere cambios iberadas deste la hipdfisis anterior en respuesta a it
sino que también las céluas folieulares que larodean _ormana liberadora de gonadotrofinas (GriRH) desde
sedividen ydan lugara un proceso denominado foli- 1 hipotélamo, Un aumento en la cemcentracién de PSH
plasmética genera reclutamien-
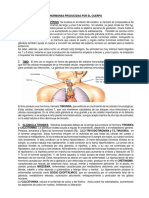
Ovogénesis y desarrollo folicalar en boyinos toperiédico de unconjunto de
iturin 1 Blteo Recital y te es lee re
Germinal de las ovogonias —meidtico —_—erecimiento ovocitario ei =
Primordial por mite oveltaio esperan dos caminos posibles:
t biotin sree oar
+O +a > “4 > foliculo dominante eventual-
270 *0+ 0-O-@ a
bordinados involucionarén y
Forete lieu tecanie ‘entrarén en atresia, Elrwimero
Oreck Primaria ‘ent ‘SsfelGllosque berate
Piimerotinetm | omoee | oesti0 esos] 152] 22 | nancia es especie-espectfico
Diimetovecrio (mm) | 20 0 8.10 |ton)noi~ | + Comoconsecuencia de un au-
x | evsoven 5 + | + [+ | mento enlaconcentracisn de
ove ose _ : : + | + | +} estradiol sintetizado desde el
Nooo + | + | folfcuto dominante, se postu-
Conscid de dvalle ¥ = +} laquetos folteulos subordina-
dos quedian expuestos a bajas
Figure 1 Representacnesquamitia dela ovine a clone en bovinos. A Congentraciones de FSH tne
medida que auments ol ditmero ovoctaro y fcr, aumenta I adguscion de ls {or Waahoreice dor ee
compotencia meidtice la epacad de desarollo. +, ocurre; no ocurm; GV, veeeula ©"
germinal GVBD, rupture de la vesieula germinal; MI'y MIL metafaws ly I, respecivae _Mientosintetizados por el fli-
mente (Rodiiguee y Farin, 2008), culo dominante,
‘ASC, CEGYR - REVISTA DE MEDICINA REPRODUCTIVA 118
Durante el perfodo de crecimiento, e1 ovocito expe-
rimenta un aumento en la tasa metabilica, lasintesis,
de ARN y proteinas, cambios en su tamafio y arqui-
tectura, y aparici6n de nuevas estructuras como la
zona peliicida y los grénulos corticales (GCs). Con-
comitante con estos cambios, adquiere progresiva-
‘mente la competencia molecular que le permitiré ini-
ciarla primera divisién meitica, progresar hacia la
segunda y detenerse en metafase para posteriormen-
te poderser receptivoa la fecundacién.
Finalmente, durante el tercer perfoda de la
ovogénesis, enominado maduracién, se pone de ma-
nifiesto la competencia meistica y se alcanza el maxi-
‘mo potencial de desarrollo embrionario. La capacidad.
de un ovocito de madurar y de dar origen a un em-
brisn depende dela acumulacién de ARN mensajeros
(ARNm) y proteinas especificas Sirard et al, 2000) las,
cuales se almacenan progresivamente durante el ceci-
rmiento y Ta maduracién ovecitaria (pertodos segundo y
tercero,respectivamente) (Wickramasinghe y Albertini,
1993). En el momento.en que se produce la ovulacién,
elovocitomaduro debe contener grupos espectficos de
proteinas y ARNm que controlaran el desarrollo em-
brionario temprano, incluyendo todas las enzimas re-
queridas parael metabolismo ovocitario.
No todas los ARNm que se encuentran en el cito-
pplasma ovocitario son apios para ser traduciclos. Exis-
te una fina regulacién dada por un proceso denomi-
nado poliadenilacin, ademés de fosforilacién de facto-
res iniciadores y proteinas ribosémicas (Tomek et al,
2001). La elongacién de la cola de poli-A de los
transcriptos se relaciona con la activaci6n de los mis-
‘mos para la traduccién mientras que el acortamiento
delacola de poli se relaciona con el silenciamiento
de estos transcriptos. De esta forma, se permite la tra-
duceién de transcriptos de manera selectiva durante
Tamaduraci6n, la fecundacién y el desarrollo embrio~
nario temprano hasta la transicién matemo-cigética,
momento en el cual se activa el genoma embrionario
(Mimeli y First, 1999).
MADURACION DE OVOCITOS: DIFERENCIAS Y
SIMILITUDES ENTRE MODELOS ANIMALES
Para completar la maduraci6n in vitro de manera
adecuada y completa, las ovocitos deben provenir de
folfculos quehayan alcanzado un tamariomninimo, tan-
toenbovines (Pavlok tal, 1992), ovinos (Szollosietal.,
1988), caprinos (Crozet etal, 1995),porcinos (Motlik et
al, 1984), monos (Schramm et al., 1993; Gilchrist etal,
11995), equinos (Goudet et al, 1997) y humanos (Tsujiet
al, 1985). La maduracién in vitrode ovecitos provenien-
tes de foliculos preantrales seguida de fecundacicn y
nacimientos fue exitosa tinicamenteen ratones. Las di-
ferencias fisiol6gicas entre especies poliovulares como
elratén y las monovulares como las especies domésti-
‘cas elhumano definen cudlesson losmejoresmodelos
animales que se pueden aproximar al estucio de los
;procesos ovocitarios en el humano.
Para un mayor entendimiento de las diferencias
entre las distintas especies de mamiferos, la siguiente
tabla muestra algunas parémetros a tener en cuenta
(ver Tabla.
El proceso de maduracién ovocitaria involucra una
serie de eventos estructurales y moleculares, que con-
llevana lamadurez nuclear y citoplasmatica y es es-
trictamente dependiente del ambiente folicular.
Los eventos relacionados a la madurez nuclear
comienzan con la ruptura de la vesfcula germinal
(Germinal Vesicle Break Down, GVBD), seguida de
condensacién cromosémica y formacién del huso
meistico, expulsién del primer cuerpo polar y final-
mente arresto de a divisin en metafase de la segur-
da division meitica (Mott y Fulks, 1976). Mientras
que la maduracién citoplasmatica involucra la loca-
Iizacién de los grénulos corticales subyacentes a la
‘membrana plasmatica en el oolema, la reorganiza-
cin de la corteza y el citoesqueleto asociado, la
rredistribucion del sistema de endomembranas (ret
culo endosplasmico y aparato de Golgi) y re-
localizacion de ARNm corticales (Sardet etal, 2002).
‘Tabla I. Algunos parémetros de ovocitos y embriones de ratdn, bovinos y humanos (de Betteridge y Rieger, 1993).
Humano Bovino Raton
Digmetro ovocitario (um) 120-150 150-180 90-100
‘Tiempo (hs.) para aleanzar:
Estadio de2células 20 %6 2
Blastodsto 120 150 ”
Eclosion, 150 200 100
Estadio de activacién del genoma embrionario | 4céiulas Scélulas 2edlulas
6
SECC. ARI. ORIG. © DE REVISION - Vol
BNP 2 Agosto 2005
MADURACION NUCLEAR.
Elreinicio de la meiosis in vivo sucede en respues-
taa la estimulacién por un pico endégeno de LH so-
bree foliculo preovulatorio, in vitro la remocién del
complejo ovocito-células del ctimulus (COC) del am-
biente folicular reinicia espontaneamente la meiosis
de aquellos ovocitos meidticamente competentes. Este
reinicio espontaneo de la meiosis se debe a que el
ambiente folicular contiene factores inhibidores de la
sis tales como el inhibidor meistico ovacitario
ipoxantinay AMP cictico (Taieb etal., 1997),
Este reinicio también puede ser inducido por
gonadotrofinas (FSH y LH) in vitro en respuesta a la
sehal desencadenada por la unién de estas hormo-
nas a sus receptores en las células del cimulus
(Rodriguez y Farin, 2004).
Lo anteriormente dicho se puede resumir en la
Figura 2,
En general, el ciclo celular de las células euicariotas
«esta regulado por dos familias de proteinas: la prime-
725 la familia de las proteinas Kinasas dependien-
tes de ciclina (Cae), las cuales inducen procesos
fosforiland blancos especificos sobre serinas y
treoninas. La segunda es la familia de proteinas
activadoras especializadas, las ciclinas que se unen
a las Cae y controlan su capacidad para fosforilar a
las protefnas blanco. El ensamblaje de ciclinas y Ce
asicomo su activacisn y desensamblaje son procesos
centrales que ditigen el ciclo celular. Uno de los prin-
También podría gustarte
- Proceso y La Instrumentación Del Proceso de Flotación de Una PDFDocumento12 páginasProceso y La Instrumentación Del Proceso de Flotación de Una PDFJhan Curay Quevedo40% (5)
- Semana 11 Triasico y Jurasico en El PerúDocumento3 páginasSemana 11 Triasico y Jurasico en El PerúDicxon DíazAún no hay calificaciones
- Reporte Cromosoma ''Y'' en Raíz de CabelloDocumento2 páginasReporte Cromosoma ''Y'' en Raíz de CabelloGladis Aimé Hernández DuránAún no hay calificaciones
- Practica 2Documento16 páginasPractica 2Andrés SerranoAún no hay calificaciones
- Programa-Procesamiento Del Semen y Selección Espermática en Reproducción AsistidaDocumento2 páginasPrograma-Procesamiento Del Semen y Selección Espermática en Reproducción AsistidaNatalia StorkAún no hay calificaciones
- Taller Hormonas Producidas Por El Cuerpo - Grado 11Documento4 páginasTaller Hormonas Producidas Por El Cuerpo - Grado 11Oscar Yamid JovenAún no hay calificaciones
- Tutoriaaa Tarea CompletaDocumento7 páginasTutoriaaa Tarea Completapstricio choquecotaAún no hay calificaciones
- Barbera Lofoscopia G1 PDFDocumento34 páginasBarbera Lofoscopia G1 PDFDUNNE3333Aún no hay calificaciones
- Un Modelo de Etapas de Desarrollo UrbanoDocumento4 páginasUn Modelo de Etapas de Desarrollo UrbanoCharliie JiimenezAún no hay calificaciones
- ) Historia de La Fitopatologia 1ij21Documento8 páginas) Historia de La Fitopatologia 1ij21David Aguilar EcheverriaAún no hay calificaciones
- CATEDRA de HistologiaDocumento136 páginasCATEDRA de HistologiaJose MedinaAún no hay calificaciones
- Guias #3 - Biologia (8) Cele y RaulDocumento12 páginasGuias #3 - Biologia (8) Cele y RaulJunior RodriguezAún no hay calificaciones
- 3 Color IMPRIMIR DEFICIENCIA DE NutricionalesDocumento6 páginas3 Color IMPRIMIR DEFICIENCIA DE Nutricionalesrafael rodriguezAún no hay calificaciones
- Preguntas Explosivas - Dinámicas CristianasDocumento1 páginaPreguntas Explosivas - Dinámicas CristianasEIMY XIOMARA TEJADA CERONAún no hay calificaciones
- Protocolo de Test Persona Bajo La Lluvia ResueltoDocumento77 páginasProtocolo de Test Persona Bajo La Lluvia ResueltoMAYRA MICHELLE ROSALES ROBLESAún no hay calificaciones
- Práctica-N02-Virologia Selección, Empleo y Esterilización Del Material en El Laboratorio Virología. Utilización de FiltrosDocumento10 páginasPráctica-N02-Virologia Selección, Empleo y Esterilización Del Material en El Laboratorio Virología. Utilización de FiltrosCarlos Moreno TafurAún no hay calificaciones
- Anatomia Del OjoDocumento8 páginasAnatomia Del OjofrankcaroAún no hay calificaciones
- Evaluacion Diagnostica 1 GradoDocumento2 páginasEvaluacion Diagnostica 1 GradoMartín Leonardo Lacuta Vara100% (1)
- Diapositivas de Los CrustaceosDocumento21 páginasDiapositivas de Los CrustaceosPilar PasaperaAún no hay calificaciones
- Neuropsicologia de Las Funciones EjecutivasDocumento104 páginasNeuropsicologia de Las Funciones EjecutivasPaula San MillánAún no hay calificaciones
- Proyecto GuasacacaDocumento27 páginasProyecto GuasacacaMerbec BaudinoAún no hay calificaciones
- Aislamiento de Enterobacterias en Lactuca Sativa, LechugaDocumento19 páginasAislamiento de Enterobacterias en Lactuca Sativa, LechugaCésar BermúdezAún no hay calificaciones
- Pajuro PDFDocumento4 páginasPajuro PDFejnavarroc100% (5)
- Atención Inmediata Del Recién NacidoDocumento18 páginasAtención Inmediata Del Recién NacidoAngelVictorioAún no hay calificaciones
- Agentes Quimicos de Irrigacion Del Sistema de Conductos RadicularesDocumento20 páginasAgentes Quimicos de Irrigacion Del Sistema de Conductos Radiculareslaura marchenaAún no hay calificaciones
- Practica 1 - Quimiotaxis BacterianaDocumento7 páginasPractica 1 - Quimiotaxis BacterianaAngelica RojasAún no hay calificaciones
- Guia TP IntroductorioDocumento2 páginasGuia TP IntroductorioDaniel MarianoAún no hay calificaciones
- Informe de MicorrizasDocumento19 páginasInforme de MicorrizasKeiv P. DavilaAún no hay calificaciones
- Mapa Conceptual Plantilla 01Documento3 páginasMapa Conceptual Plantilla 01MELISSA ESTRADA VALLADOLIDAún no hay calificaciones
- Tema07 BernoulliDocumento18 páginasTema07 BernoulliJose Luis Rodríguez DíazAún no hay calificaciones