También podría gustarte
- Precursores y macromoléculas biológicasDocumento31 páginasPrecursores y macromoléculas biológicasElisa nin EncarnacionAún no hay calificaciones
- Cáncer de MamaDocumento16 páginasCáncer de MamaElisa nin EncarnacionAún no hay calificaciones
- Estudio Comparativo de Los Precursores de Las Macromoleculas y Las Macromoleculas (2) Noelmi 1Documento2 páginasEstudio Comparativo de Los Precursores de Las Macromoleculas y Las Macromoleculas (2) Noelmi 1Elisa nin EncarnacionAún no hay calificaciones
- Accidente Cerebro Va 103094 Downloadable 524799Documento13 páginasAccidente Cerebro Va 103094 Downloadable 524799Elisa nin EncarnacionAún no hay calificaciones
- Caso Clinico de LipomaDocumento21 páginasCaso Clinico de LipomaElisa nin EncarnacionAún no hay calificaciones
- Tema 31 ToxicologiaDocumento18 páginasTema 31 ToxicologiaElisa nin EncarnacionAún no hay calificaciones
- MIP-133 Pandemia 2020 Unidad 2Documento113 páginasMIP-133 Pandemia 2020 Unidad 2Elisa nin EncarnacionAún no hay calificaciones
- Cuestionario Guia Acidoo BaseDocumento4 páginasCuestionario Guia Acidoo BaseElisa nin EncarnacionAún no hay calificaciones
- Giardiosis: parásito intestinal Giardia lambliaDocumento6 páginasGiardiosis: parásito intestinal Giardia lambliaYohanly Hernández AlmonteAún no hay calificaciones
- Adenocarcinomas Cindy de Los SantosDocumento10 páginasAdenocarcinomas Cindy de Los SantosElisa nin EncarnacionAún no hay calificaciones
- PBO2 CNNP PresentaciónDocumento18 páginasPBO2 CNNP PresentaciónElisa nin EncarnacionAún no hay calificaciones
- Tema 2 Marco LegalDocumento10 páginasTema 2 Marco LegalElisa nin EncarnacionAún no hay calificaciones
- AlcanosDocumento22 páginasAlcanosGabriel Jaquez FortunaAún no hay calificaciones
- Apuntes - Clases DermatoDocumento49 páginasApuntes - Clases DermatoLuis SilverioAún no hay calificaciones
- Cfi-122 Genetica MedicaDocumento15 páginasCfi-122 Genetica Medicafranklin medranoAún no hay calificaciones
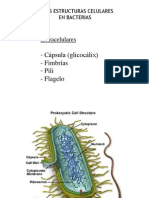
- Estructura BacterianaDocumento4 páginasEstructura BacterianaElisa nin EncarnacionAún no hay calificaciones
- Actividad Herencia MultifactorialDocumento4 páginasActividad Herencia MultifactorialElisa nin EncarnacionAún no hay calificaciones
- Potencial de AccionDocumento19 páginasPotencial de AccionElisa nin EncarnacionAún no hay calificaciones
- Tema 17 Medicina Legal AntropologicaDocumento28 páginasTema 17 Medicina Legal AntropologicaElisa nin EncarnacionAún no hay calificaciones
- PBO2 CNNP PresentaciónDocumento18 páginasPBO2 CNNP PresentaciónElisa nin EncarnacionAún no hay calificaciones
- Biocatalizador Virtual 2020-2Documento58 páginasBiocatalizador Virtual 2020-2Elisa nin EncarnacionAún no hay calificaciones
- Libro Microbiología Básica para El Área de La Salud y Afines (HUGO)Documento244 páginasLibro Microbiología Básica para El Área de La Salud y Afines (HUGO)ivan100% (1)
- Semiología QuirúrgicaDocumento102 páginasSemiología QuirúrgicaDanielaAún no hay calificaciones
- Giardiosis: parásito intestinal Giardia lambliaDocumento6 páginasGiardiosis: parásito intestinal Giardia lambliaYohanly Hernández AlmonteAún no hay calificaciones
- Ashwagandha Natura FoundationDocumento32 páginasAshwagandha Natura FoundationJean TejeraAún no hay calificaciones
- Pattern Recognition Receptors (Receptores Que Reconocen Patrones (PRRS) )Documento4 páginasPattern Recognition Receptors (Receptores Que Reconocen Patrones (PRRS) )Paul Suarez MaldonadoAún no hay calificaciones
- Unidad 3 - Sistemas de EndomembranasDocumento17 páginasUnidad 3 - Sistemas de EndomembranasMelAún no hay calificaciones
- Cuadro Comparativo de Desarrollo de Linaje HematopoyeticoDocumento5 páginasCuadro Comparativo de Desarrollo de Linaje Hematopoyeticopaot27Aún no hay calificaciones
- Sist GustativoDocumento44 páginasSist GustativoDaniel RamírezAún no hay calificaciones
- Archivo 2 Bases Moleculares Del Desarrollo EmbrionarioDocumento36 páginasArchivo 2 Bases Moleculares Del Desarrollo EmbrionarioYar CBAún no hay calificaciones
- Caspasas: mediadores apoptosisDocumento4 páginasCaspasas: mediadores apoptosisJoselyn KarinaAún no hay calificaciones
- Señalización CelularDocumento32 páginasSeñalización CelularCamilo Andrés RuedaAún no hay calificaciones
- HongosDocumento65 páginasHongosJose Manuel Morillo RodriguezAún no hay calificaciones
- Arriaga González, I. (2021)Documento74 páginasArriaga González, I. (2021)Junior CordovaAún no hay calificaciones
- ASEA REDOX Folleto ESDocumento16 páginasASEA REDOX Folleto ESCanon TransparenteAún no hay calificaciones
- Conceptos Básicos de La Señalización Molecular en El Desarrollo PDFDocumento11 páginasConceptos Básicos de La Señalización Molecular en El Desarrollo PDFDana P Rosado TrujilloAún no hay calificaciones
- Señalización Celular, Camilo Escobedo Fernández PDFDocumento5 páginasSeñalización Celular, Camilo Escobedo Fernández PDFcamiloesferAún no hay calificaciones
- Fisiopatología de la diabetes tipo 2: resistencia a la insulina y disfunción de las células βDocumento5 páginasFisiopatología de la diabetes tipo 2: resistencia a la insulina y disfunción de las células βrj1406Aún no hay calificaciones
- Genetica 8° ClaseDocumento42 páginasGenetica 8° ClaseIngrid BGAún no hay calificaciones
- Receptores para Farmacos, y FarmacodinamiaDocumento41 páginasReceptores para Farmacos, y FarmacodinamiaMarilyn DardonAún no hay calificaciones
- AntecedentesDocumento10 páginasAntecedentesDavid GuzmanAún no hay calificaciones
- Discusion 1. Bqm2-1Documento29 páginasDiscusion 1. Bqm2-1Verónica Michelle Beltrán VelisAún no hay calificaciones
- Clase 5 RENOVACION, REPARACION Y REGENERACION TISULAR Clase Virtual 2022Documento44 páginasClase 5 RENOVACION, REPARACION Y REGENERACION TISULAR Clase Virtual 2022persy dafneAún no hay calificaciones
- Relación Dosis-Respuesta Gradual de Acetilcolina, Carbacol y PilocarpinaDocumento6 páginasRelación Dosis-Respuesta Gradual de Acetilcolina, Carbacol y PilocarpinaKarla Cabrera100% (1)
- Seminario Receptores UPAODocumento21 páginasSeminario Receptores UPAODiego GutierrezAún no hay calificaciones
- Clase 4 Capsula, Frimbia, Pili, FlageloDocumento74 páginasClase 4 Capsula, Frimbia, Pili, Flageloapi-19878431100% (1)
- 15.1 Los Elementos Básicos de Los Sistemas de Señalización CelularDocumento6 páginas15.1 Los Elementos Básicos de Los Sistemas de Señalización CelularALEJANDRA INÉS NIETO ARIASAún no hay calificaciones
- El Papel Del Microambiente Tumoral y Los Fibroblastos Asociados A Cáncer en El Desarrollo y La Progresión Del Cáncer de MamaDocumento12 páginasEl Papel Del Microambiente Tumoral y Los Fibroblastos Asociados A Cáncer en El Desarrollo y La Progresión Del Cáncer de MamaCynthia RamírezAún no hay calificaciones
- Taller de Repaso Bioseñalización y Bioenergética Marzo 2020Documento3 páginasTaller de Repaso Bioseñalización y Bioenergética Marzo 2020mafe ospinaAún no hay calificaciones
- Regulacion de La Exprecion Genica, Sistema InmunitarioDocumento17 páginasRegulacion de La Exprecion Genica, Sistema InmunitarioKleverxithoDiazCiezaAún no hay calificaciones
- Proteínas G: estructura, clasificación, funciones y su importancia en la transducción de señalesDocumento19 páginasProteínas G: estructura, clasificación, funciones y su importancia en la transducción de señalesEduardo P. FernándezAún no hay calificaciones
- Albumcelulaslinfoides, ViasfranklinvDocumento9 páginasAlbumcelulaslinfoides, ViasfranklinvJuan BanegasAún no hay calificaciones
- Temario PUEM AnestesiologiaDocumento4 páginasTemario PUEM AnestesiologiaReyna67% (3)
- CitoquinasDocumento10 páginasCitoquinasValentina Robayo AguirreAún no hay calificaciones