También podría gustarte
- Bases de la electrofisiología celularDocumento9 páginasBases de la electrofisiología celularMaria Paz CUELLOAún no hay calificaciones
- Clase 4 - 5 Excitabilidad y Transmisión SinàpticaDocumento47 páginasClase 4 - 5 Excitabilidad y Transmisión SinàpticaJohana Andrea Cisterna VegaAún no hay calificaciones
- Excitabilidad CelularDocumento51 páginasExcitabilidad CelularAnonymous RbHHOl1tjAún no hay calificaciones
- Sem 3 Biofisica Fen Bioelectricos y MembranaDocumento60 páginasSem 3 Biofisica Fen Bioelectricos y MembranaCARLOS LUCIANO TORRES FERNANDEZAún no hay calificaciones
- Potencial de Membrana.Documento6 páginasPotencial de Membrana.Carlos BarriosAún no hay calificaciones
- Propiedades Eléctricas de Las Membranas de Las Células ExcitablesDocumento5 páginasPropiedades Eléctricas de Las Membranas de Las Células ExcitablesCésar Yael Partida Pérez50% (2)
- Potencial de MembranaDocumento38 páginasPotencial de MembranaWilliam MartinezAún no hay calificaciones
- Potencial de MembranaDocumento17 páginasPotencial de MembranaRobert AcostaAún no hay calificaciones
- Seminario Biopotenciales FisiologiaDocumento9 páginasSeminario Biopotenciales Fisiologiayamirjossue100% (9)
- Potencial de Membrana 1Documento17 páginasPotencial de Membrana 1academiusAún no hay calificaciones
- La Generación y La Transmisión Del Impulso NerviosoDocumento21 páginasLa Generación y La Transmisión Del Impulso NerviosomixoneqAún no hay calificaciones
- Origen de Los Biopotenciales: 86.62 Equipamiento para Diagnostico y Tratamiento Biomédico Dr. Ing. Esteban ValverdeDocumento38 páginasOrigen de Los Biopotenciales: 86.62 Equipamiento para Diagnostico y Tratamiento Biomédico Dr. Ing. Esteban ValverdeElmaster420Aún no hay calificaciones
- Potenc.y Sinap. Rhoades 5edDocumento56 páginasPotenc.y Sinap. Rhoades 5edYerlin Murillo MongeAún no hay calificaciones
- NERNSTDocumento5 páginasNERNSTleslieAún no hay calificaciones
- Clase 1 Biol 178 20212Documento35 páginasClase 1 Biol 178 20212Estefania conejerosAún no hay calificaciones
- Potenciales de membrana en células nerviosas y muscularesDocumento17 páginasPotenciales de membrana en células nerviosas y musculareslilia estefania B BalarezoAún no hay calificaciones
- Canales Iónicos y Potenciales MembranaDocumento17 páginasCanales Iónicos y Potenciales MembranaAnthony Namay PoloAún no hay calificaciones
- Bases Biofisicas de Fenomenos EléctricosDocumento14 páginasBases Biofisicas de Fenomenos EléctricosDiego Sanchez PerezAún no hay calificaciones
- Elementos celulares del SNC y excitabilidad celularDocumento32 páginasElementos celulares del SNC y excitabilidad celularSirwinston Churchill Jr.Aún no hay calificaciones
- Transmisión señales sistema nerviosoDocumento13 páginasTransmisión señales sistema nerviososilverwolf_aqpAún no hay calificaciones
- Potencial de MembranaDocumento29 páginasPotencial de MembranaMaria FernandaAún no hay calificaciones
- Ecuacion de NerstDocumento8 páginasEcuacion de NerstIvonne Andrea Donoso ToledoAún no hay calificaciones
- Laboratorio Potencial de Membrana 1Documento5 páginasLaboratorio Potencial de Membrana 1Luis Eduardo GutierrezAún no hay calificaciones
- Unidad 4Documento3 páginasUnidad 4J . Arturo Corrales HernándezAún no hay calificaciones
- TEJIDOS EXCITABLES Guia de Estudio PDFDocumento42 páginasTEJIDOS EXCITABLES Guia de Estudio PDFMaria Luz VirasoroAún no hay calificaciones
- Actividad Practica 3 - Biomembrana FBEKyF 2023Documento4 páginasActividad Practica 3 - Biomembrana FBEKyF 2023lolaAún no hay calificaciones
- Neurofisiología: Clase 2Documento43 páginasNeurofisiología: Clase 2Ian Paulo Villagran LeonAún no hay calificaciones
- 3 Origen Del Potencial de MembranaDocumento20 páginas3 Origen Del Potencial de MembranaAlan PardaveAún no hay calificaciones
- Potenciales de MembranaDocumento36 páginasPotenciales de Membranapepe_074Aún no hay calificaciones
- BIOPOTENCIALES FisiologíaDocumento6 páginasBIOPOTENCIALES FisiologíaDJ EDWAún no hay calificaciones
- Cuarta Semana BioDocumento50 páginasCuarta Semana Biomike kaiser romanovAún no hay calificaciones
- ElectrofiosiologíaDocumento3 páginasElectrofiosiologíaEduardo Jahir AnguloAún no hay calificaciones
- Potencial de MembranaDocumento3 páginasPotencial de MembranaJoel DuarteAún no hay calificaciones
- 2.1potenciales de MembranaDocumento175 páginas2.1potenciales de MembranaMARIANA GOMEZ CHAVEZAún no hay calificaciones
- Bioelectricidad Introducción PDFDocumento43 páginasBioelectricidad Introducción PDFPamela Liendo MartinezAún no hay calificaciones
- Bioelectric I DadDocumento126 páginasBioelectric I DadJean VillanuevaAún no hay calificaciones
- FisiologíaDocumento4 páginasFisiologíaMartha PadillaAún no hay calificaciones
- Excitabilidad Celular - BiofísicaDocumento48 páginasExcitabilidad Celular - BiofísicaChucho Reach100% (3)
- Tema 3. Potencial de MembranaDocumento11 páginasTema 3. Potencial de MembranaAlbaAún no hay calificaciones
- ELECTROFISIOLOGIADocumento68 páginasELECTROFISIOLOGIAaleciram_goodface227100% (3)
- Tejido ExcitableDocumento90 páginasTejido ExcitablepierregggAún no hay calificaciones
- TP 4 PropiedadespasivasdelamembranaDocumento10 páginasTP 4 PropiedadespasivasdelamembranaVero OrtegaAún no hay calificaciones
- Act. BiomembranasDocumento7 páginasAct. BiomembranasGiannina GattiAún no hay calificaciones
- BIOELECTROMAGNETISMODocumento96 páginasBIOELECTROMAGNETISMOFátima Arias CabrejosAún no hay calificaciones
- BiopotencialesDocumento13 páginasBiopotencialesWanda Andrade CastroAún no hay calificaciones
- Tema 3. Equilibrio Ionico y Potencial de Membrana de ReposoDocumento26 páginasTema 3. Equilibrio Ionico y Potencial de Membrana de ReposoXimenna Antuaneth TapiaAún no hay calificaciones
- Potenciales membrana y transporte iónicoDocumento62 páginasPotenciales membrana y transporte iónicoJuan Pablo Díaz BecerraAún no hay calificaciones
- Potencial de Accion y MembranaDocumento8 páginasPotencial de Accion y MembranaFrancisco MoroyoquiAún no hay calificaciones
- B 2 BacaDocumento32 páginasB 2 Bacajgfb180165Aún no hay calificaciones
- Neurofisiología Rev PDFDocumento41 páginasNeurofisiología Rev PDFSergio Oriol Macias HernandezAún no hay calificaciones
- NeurofisiologiaDocumento10 páginasNeurofisiologiaDANIEL ALEJANDROAún no hay calificaciones
- Potencial de MembranaDocumento12 páginasPotencial de Membranadanitza pilcoAún no hay calificaciones
- Potencial de NernstDocumento4 páginasPotencial de Nernsta7024782Aún no hay calificaciones
- 2 (1) (1) ClasePotencialdeMembranaDocumento21 páginas2 (1) (1) ClasePotencialdeMembranaBastian Rodrigo Almendra FernandezAún no hay calificaciones
- Biofísica Del Sistema NerviosoDocumento30 páginasBiofísica Del Sistema NerviosoNafe Lopez67% (3)
- Conducta Eléctrica de La MembranaDocumento68 páginasConducta Eléctrica de La Membranajuarddan100% (1)
- Experiencias sobre circuitos eléctricos en serie operados por corriente directaDe EverandExperiencias sobre circuitos eléctricos en serie operados por corriente directaAún no hay calificaciones
- Competencias de EmpleabilidadDocumento17 páginasCompetencias de EmpleabilidadGerardo Antonio Cornejo BallesterosAún no hay calificaciones
- PsicodiagnósticoDocumento63 páginasPsicodiagnósticoJulia ScavoneAún no hay calificaciones
- Desarrollo Del Sistema LocomotorDocumento56 páginasDesarrollo Del Sistema LocomotorNicole GuillénAún no hay calificaciones
- PulpectomíaendientestemporalesDocumento86 páginasPulpectomíaendientestemporalesSylvia Tonantzin Diaz VegaAún no hay calificaciones
- Diapos Dr. Vargas RespiratorioDocumento133 páginasDiapos Dr. Vargas RespiratorioOscar David Tecse OrtizAún no hay calificaciones
- Meditacion ZenDocumento3 páginasMeditacion ZenAntuanoNoventaAún no hay calificaciones
- Arrugas de La PielDocumento5 páginasArrugas de La PielHugo Villca LutinoAún no hay calificaciones
- Operaciones Ampliadoras Del Canal de PartoDocumento20 páginasOperaciones Ampliadoras Del Canal de PartoDaniel Ozuna Gordillo100% (2)
- Sistema de las cininas y coagulaciónDocumento12 páginasSistema de las cininas y coagulacióncarmen gonzalezAún no hay calificaciones
- Tubo digestivo IIDocumento9 páginasTubo digestivo IIRegina Cortés100% (1)
- Alfa AmilasaDocumento1 páginaAlfa AmilasaAldhair Moreno JassoAún no hay calificaciones
- Seminario 3 GlucolisisDocumento20 páginasSeminario 3 GlucolisisJosé MoralesAún no hay calificaciones
- Accesos Venosos EcoguiadosDocumento22 páginasAccesos Venosos EcoguiadosVlady78Aún no hay calificaciones
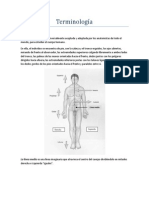
- Terminología Educación para La SaludDocumento8 páginasTerminología Educación para La SaludNatalia Sánchez100% (2)
- Existen 3 Frenillos Labial SuperiorDocumento9 páginasExisten 3 Frenillos Labial SuperiorBeto NavaAún no hay calificaciones
- Fenómenos ContráctilesDocumento7 páginasFenómenos ContráctilesJosé MoralesAún no hay calificaciones
- Evaluación Órganos de FonoarticulatoriosDocumento6 páginasEvaluación Órganos de FonoarticulatoriosFernando Ayala SandovalAún no hay calificaciones
- Sexual I DadDocumento52 páginasSexual I DadAdalberto López SilvaAún no hay calificaciones
- Alimentos para La Concentracion y MemoriaDocumento5 páginasAlimentos para La Concentracion y MemoriaSusy LopezAún no hay calificaciones
- Tesis Publicidad SubliminalDocumento421 páginasTesis Publicidad SubliminalHolman Delgado CAún no hay calificaciones
- Caso ClínicoDocumento41 páginasCaso Clínicotasho1998Aún no hay calificaciones
- Graficas Estrés AcadémicoDocumento18 páginasGraficas Estrés AcadémicoAnye Arteaga MiramónAún no hay calificaciones
- Ficha de PeneDocumento4 páginasFicha de PeneKaren GutierrezAún no hay calificaciones
- Hipoplasia Medular ClaseDocumento46 páginasHipoplasia Medular ClaseSharom R. AlvarezAún no hay calificaciones
- Ficha Sistema Nervioso Periferico para Sexto de PrimariaDocumento5 páginasFicha Sistema Nervioso Periferico para Sexto de Primarianatividad ordoño alca100% (2)
- Análisis Transversal de ModelosDocumento13 páginasAnálisis Transversal de ModelosSelene RamosAún no hay calificaciones
- Hidroterapia, agua y saludDocumento18 páginasHidroterapia, agua y saludluis3107Aún no hay calificaciones
- Tesis de Trabajo de Aban Dono Micronutrientes (Autoguardado)Documento35 páginasTesis de Trabajo de Aban Dono Micronutrientes (Autoguardado)Víctor Amador Bravo Cauna50% (2)
- SPOROZOARIOSDocumento8 páginasSPOROZOARIOSDiego MuñozAún no hay calificaciones
- 5 Diagramas de Flujo Signos VitalesDocumento17 páginas5 Diagramas de Flujo Signos VitalesAnahiSanchez33% (3)