También podría gustarte
- Practica 1 EnzimasDocumento15 páginasPractica 1 EnzimasSamantha0523Aún no hay calificaciones
- Estructura y funciones de las proteínasDocumento63 páginasEstructura y funciones de las proteínasAnghela Aquino RiosAún no hay calificaciones
- LipidosDocumento61 páginasLipidosRodrigo Arce100% (1)
- Célula VegetalDocumento6 páginasCélula VegetalCamila Traslaviña100% (1)
- Estructura y enfermedades de los virusDocumento9 páginasEstructura y enfermedades de los virusHenry NoveraAún no hay calificaciones
- BasesNitrogenadas PDFDocumento47 páginasBasesNitrogenadas PDFAlfonso Martínez BravoAún no hay calificaciones
- Clasificacion de Productos Naturales Basado en La BiogenesisDocumento3 páginasClasificacion de Productos Naturales Basado en La BiogenesisJosué AduvireAún no hay calificaciones
- Bioquimica 1. Analisis Cuantitativo de EnzimasDocumento12 páginasBioquimica 1. Analisis Cuantitativo de EnzimasangelaAún no hay calificaciones
- Identificación carbohidratosDocumento26 páginasIdentificación carbohidratosAmada100% (1)
- CerebrosidosDocumento2 páginasCerebrosidosJose Gregorio Davila HernandezAún no hay calificaciones
- Digestión y absorción de hidratos de carbonoDocumento7 páginasDigestión y absorción de hidratos de carbonoSTUDY ACCOUNTAún no hay calificaciones
- Ensayo Envejecimiento y Muerte CelularDocumento14 páginasEnsayo Envejecimiento y Muerte CelularLambda GarcíaAún no hay calificaciones
- Importancia de La Bioquimica en Ipq-2019 PDFDocumento151 páginasImportancia de La Bioquimica en Ipq-2019 PDFMarlon FuentesAún no hay calificaciones
- Informe de Laboratorio: Reconocimiento de BiomoléculasDocumento24 páginasInforme de Laboratorio: Reconocimiento de BiomoléculasAndresAún no hay calificaciones
- IntroducciónDocumento5 páginasIntroducciónFabián AndrésAún no hay calificaciones
- Influencia de La Saccharomyces Cerevisiae en El PanDocumento40 páginasInfluencia de La Saccharomyces Cerevisiae en El PanfloriabaAún no hay calificaciones
- LipidosDocumento51 páginasLipidosKimberly Sanchez Sanchez .100% (1)
- Proteinas y EnzimasDocumento48 páginasProteinas y EnzimasWillSmith SanchezAún no hay calificaciones
- Acidos NucleicosDocumento37 páginasAcidos NucleicosBrenda YepesAún no hay calificaciones
- Práctico ProteínasDocumento3 páginasPráctico ProteínasVeronica Rodriguez MoyanoAún no hay calificaciones
- Esteroles (Colesterol)Documento8 páginasEsteroles (Colesterol)Brando Molina NuñezAún no hay calificaciones
- Nucleotidos Bases Puricas y Pirimidicas1Documento13 páginasNucleotidos Bases Puricas y Pirimidicas1lopz_catherineAún no hay calificaciones
- Ácidos nucleicos: estructura y funcionesDocumento20 páginasÁcidos nucleicos: estructura y funcionesSherly Yesenia Melgarejo CastañedaAún no hay calificaciones
- INFORMEDocumento10 páginasINFORMEAnders GarcíaAún no hay calificaciones
- Informe de Laboratorio No. 3 Identificación de LipidosDocumento10 páginasInforme de Laboratorio No. 3 Identificación de LipidosKatherin BernalAún no hay calificaciones
- GlicocalixDocumento12 páginasGlicocalixTeo NavarroAún no hay calificaciones
- Protozoarios 1Documento19 páginasProtozoarios 1Inaoris MdoAún no hay calificaciones
- Practica 4Documento5 páginasPractica 4Luisa OrtizAún no hay calificaciones
- VitaminasDocumento57 páginasVitaminasAlvaro Josue Mazariegos FarfanAún no hay calificaciones
- Acidos Nucleicos 17-18-170910104832Documento17 páginasAcidos Nucleicos 17-18-170910104832Anonymous gguTkwGAAún no hay calificaciones
- Monografia AminoacidosDocumento15 páginasMonografia Aminoacidosjuan francisco100% (1)
- La Organización Celular y El MetabolismoDocumento21 páginasLa Organización Celular y El MetabolismoCristhian RodríguezAún no hay calificaciones
- Estructura y composición de los ácidos nucleicos (ADN y ARNDocumento23 páginasEstructura y composición de los ácidos nucleicos (ADN y ARNDalia MonteagudoAún no hay calificaciones
- 3.esterilización y DesinfecciónDocumento37 páginas3.esterilización y Desinfecciónanthony100% (1)
- Informe de Electroforesis de ProteinasDocumento15 páginasInforme de Electroforesis de ProteinasNicolas Sanchez GomezAún no hay calificaciones
- Fermentación y RespiraciónDocumento15 páginasFermentación y RespiraciónMariaAún no hay calificaciones
- Tejidos VegetalesDocumento63 páginasTejidos VegetalesGary BrianAún no hay calificaciones
- T 14 A Oxido Reduccion BiologicaDocumento19 páginasT 14 A Oxido Reduccion BiologicaElias Del aguila Perez100% (1)
- 1.introduccion Al MetabolismoDocumento9 páginas1.introduccion Al MetabolismoAndres Felipe Prieto AlarconAún no hay calificaciones
- Vitaminas BioquimicaDocumento6 páginasVitaminas BioquimicaMonika Etienne Notario100% (1)
- FOSFOLIPIDOSDocumento6 páginasFOSFOLIPIDOSalis castilloAún no hay calificaciones
- MACROELEMENTOSDocumento10 páginasMACROELEMENTOSVictor SalazarAún no hay calificaciones
- Proenzimas e IsoenzimasDocumento7 páginasProenzimas e IsoenzimasJonathan Fontana Quiroz50% (2)
- 15 Preguntas LipidosDocumento5 páginas15 Preguntas LipidosKassandra VegaAún no hay calificaciones
- Informe de Membranas CelularesDocumento6 páginasInforme de Membranas CelularesAlejandra Bravo Iriarte100% (3)
- ALMIDONDocumento24 páginasALMIDONJohacvAún no hay calificaciones
- PROTEINAS Exposicion Power PointDocumento29 páginasPROTEINAS Exposicion Power PointRommel Rodriguez100% (9)
- Resumen - Biologia ADN y ARNDocumento2 páginasResumen - Biologia ADN y ARNDaniela ChirinosAún no hay calificaciones
- LanzaderasDocumento20 páginasLanzaderasAdrian Aguirre AlvaAún no hay calificaciones
- Anexo No 2 Modelos AtómicosDocumento4 páginasAnexo No 2 Modelos AtómicosLuz Nelly Maya OrtizAún no hay calificaciones
- Informe Aminoacidos y ProteinasDocumento7 páginasInforme Aminoacidos y ProteinasLyseth GómezAún no hay calificaciones
- Tipos de MacromoléculasDocumento6 páginasTipos de Macromoléculasfeiita100% (2)
- La AmilasDocumento9 páginasLa Amilastania liset100% (1)
- Biofísica lípidosDocumento4 páginasBiofísica lípidosSamuel Aldair50% (2)
- Principios de La Lógica Molecular y Jerarquía de La OrganizacionDocumento28 páginasPrincipios de La Lógica Molecular y Jerarquía de La OrganizacionAlexanderSKateAún no hay calificaciones
- CARBOHIDRATOSDocumento52 páginasCARBOHIDRATOSMatus Ska Auriazul100% (1)
- Cariotipo Humano-1Documento11 páginasCariotipo Humano-1Alexi SuarezAún no hay calificaciones
- Ácidos NucleicosDocumento14 páginasÁcidos Nucleicosthyago pereyraAún no hay calificaciones
- Lectura Acidos Nucleicos 2022Documento9 páginasLectura Acidos Nucleicos 2022Naomy Burgos GonzalezAún no hay calificaciones
- GuiaDocumento13 páginasGuiaMichiAún no hay calificaciones
- Capítulo 7 - Iones ComplejosDocumento25 páginasCapítulo 7 - Iones ComplejosPatricia Bustelo100% (1)
- ENOVA 6°2° PresentaciónDocumento9 páginasENOVA 6°2° PresentaciónPatricia BusteloAún no hay calificaciones
- Cap 15 Metabolismo de AminoácidosDocumento45 páginasCap 15 Metabolismo de AminoácidosPatricia BusteloAún no hay calificaciones
- Modelos AtómicosDocumento21 páginasModelos AtómicosPatricia BusteloAún no hay calificaciones
- Composicion Quimica Del PlanetaDocumento66 páginasComposicion Quimica Del PlanetaPatricia BusteloAún no hay calificaciones
- Anexo 1 Teoría Atómico-MolecularDocumento8 páginasAnexo 1 Teoría Atómico-MolecularPatricia BusteloAún no hay calificaciones
- F09. Tiro Vertical y Caída LibreDocumento3 páginasF09. Tiro Vertical y Caída LibrePatricia BusteloAún no hay calificaciones
- Composicion Quimica Del PlanetaDocumento66 páginasComposicion Quimica Del PlanetaPatricia BusteloAún no hay calificaciones
- Fórmula y Nomenclatura Compuestos InorgánicosDocumento9 páginasFórmula y Nomenclatura Compuestos InorgánicosPatricia BusteloAún no hay calificaciones
- Composicion Quimica Del PlanetaDocumento66 páginasComposicion Quimica Del PlanetaPatricia BusteloAún no hay calificaciones
- Capítulo 1 - Química Del CarbonoDocumento8 páginasCapítulo 1 - Química Del CarbonoPatricia BusteloAún no hay calificaciones
- Documento de 80 líneasDocumento5 páginasDocumento de 80 líneasPatricia BusteloAún no hay calificaciones
- Maquinas SimplesDocumento53 páginasMaquinas SimplesPatricia BusteloAún no hay calificaciones
- Geografía AmbientalDocumento51 páginasGeografía AmbientalPatricia BusteloAún no hay calificaciones
- Krahg Helge Generaciones Cuanticas PDFDocumento455 páginasKrahg Helge Generaciones Cuanticas PDFPatricia Bustelo100% (1)
- RespuestasDocumento82 páginasRespuestasPatricia BusteloAún no hay calificaciones
- PLAN de CLASE (Para Completar)Documento3 páginasPLAN de CLASE (Para Completar)Patricia BusteloAún no hay calificaciones
- Propiedades ColigativasDocumento20 páginasPropiedades ColigativasPatricia BusteloAún no hay calificaciones
- Biofísica 2018Documento31 páginasBiofísica 2018Patricia BusteloAún no hay calificaciones
- Biofísica 2018Documento31 páginasBiofísica 2018Patricia BusteloAún no hay calificaciones
- Biofísica 2018Documento2 páginasBiofísica 2018Patricia BusteloAún no hay calificaciones
- Tipos de Reacciones QuímicasDocumento26 páginasTipos de Reacciones QuímicasPatricia BusteloAún no hay calificaciones
- La Corrosión Del AluminioDocumento3 páginasLa Corrosión Del AluminioPatricia BusteloAún no hay calificaciones
- Biofísica 2018Documento31 páginasBiofísica 2018Patricia BusteloAún no hay calificaciones
- Aplicación de Ondas en MedicinaDocumento15 páginasAplicación de Ondas en MedicinaPatricia BusteloAún no hay calificaciones
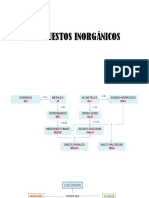
- Compuestos InorgánicosDocumento7 páginasCompuestos InorgánicosPatricia BusteloAún no hay calificaciones
- Compuestos InorgánicosDocumento10 páginasCompuestos InorgánicosPatricia BusteloAún no hay calificaciones
- Propiedades ColigativasDocumento1 páginaPropiedades ColigativasCarlosAndresBuelvasRodriguezAún no hay calificaciones
- Nivel SubmicroscópicoDocumento20 páginasNivel SubmicroscópicoPatricia Bustelo100% (1)
- TP N°-Propiedades PeriódicasDocumento2 páginasTP N°-Propiedades PeriódicasPatricia BusteloAún no hay calificaciones
- Presentacion Enlace CovalenteDocumento27 páginasPresentacion Enlace CovalentePaulaAún no hay calificaciones
- Practica 3Documento5 páginasPractica 3EmanuelGarciaAún no hay calificaciones
- Examen Química Primer Periodo Sección PreparatoriaDocumento6 páginasExamen Química Primer Periodo Sección PreparatoriaChristianAún no hay calificaciones
- Bonos Interbank US$300MMDocumento11 páginasBonos Interbank US$300MMGian Franco Quipan LayzaAún no hay calificaciones
- Criterios de Calificación Standar PoorDocumento20 páginasCriterios de Calificación Standar PoorEldyn Fernando BiggsAún no hay calificaciones
- Eteres, Esteres, Anhidridos FDocumento8 páginasEteres, Esteres, Anhidridos Fmazttr666Aún no hay calificaciones
- Problemas Resueltos de Quimica Segundo de BachilleratoDocumento68 páginasProblemas Resueltos de Quimica Segundo de Bachilleratohuaycochea100% (1)
- El Ácido SalicílicoDocumento49 páginasEl Ácido SalicílicoCristian Joaquin ZegarraAún no hay calificaciones
- Quc3admica Orgc3a1nicaDocumento86 páginasQuc3admica Orgc3a1nicaJairo Rodriguez ArgomedoAún no hay calificaciones
- Formación de osazonas, prueba de Fehling y pentaacetato de glucosaDocumento7 páginasFormación de osazonas, prueba de Fehling y pentaacetato de glucosaNayeli IbarraAún no hay calificaciones
- Solución de Laboratorio de Orgánica 1Documento1 páginaSolución de Laboratorio de Orgánica 1Mayi C LopezAún no hay calificaciones
- Estructura LewisDocumento10 páginasEstructura LewisJluissilvAún no hay calificaciones
- QIIIDocumento101 páginasQIIIBryan VargasAún no hay calificaciones
- Obtención de Anhídrido Ftálico A Partir de Acido Ftálico Con Calentamiento y Reflujo de Anhídrido AcéticoDocumento6 páginasObtención de Anhídrido Ftálico A Partir de Acido Ftálico Con Calentamiento y Reflujo de Anhídrido AcéticoHerman Ruiz CalderinAún no hay calificaciones
- Compuestos Orgánicos de AzufreDocumento20 páginasCompuestos Orgánicos de AzufreJeanneth VásconezAún no hay calificaciones
- Síntesis de benzalacetofenona en 68Documento4 páginasSíntesis de benzalacetofenona en 68Duulce AnaaiAún no hay calificaciones
- Tarea de Sopa de LetrasDocumento78 páginasTarea de Sopa de LetrasMilagros Milu MiluscaAún no hay calificaciones
- OclusiónDocumento5 páginasOclusiónEstrella MedinaAún no hay calificaciones
- Acidez y Basicidad en Productos Cotidianos (Autoguardado)Documento10 páginasAcidez y Basicidad en Productos Cotidianos (Autoguardado)jusaAún no hay calificaciones
- Proteinas ImportanteDocumento43 páginasProteinas ImportanteRuben MarquezAún no hay calificaciones
- Reporte #6Documento7 páginasReporte #6Isberto Martínez0% (1)
- Piridina (Apuntes) 30665Documento12 páginasPiridina (Apuntes) 30665bresojiarAún no hay calificaciones
- Cuaderno de Actividades Complementarias 2º Bachillerato. Química. Unidad 4.Documento4 páginasCuaderno de Actividades Complementarias 2º Bachillerato. Química. Unidad 4.Merche García SáezAún no hay calificaciones
- Taller 1 Y2Documento8 páginasTaller 1 Y2andyAún no hay calificaciones
- Derivados de Acidos CarboxilicosDocumento23 páginasDerivados de Acidos CarboxilicosYancy JuanAún no hay calificaciones
- To Del Arn de LevaduraDocumento7 páginasTo Del Arn de LevaduraGina Martinez100% (1)
- Eteres y EsteresDocumento13 páginasEteres y EsteresXimena Acevedo Moncada100% (1)
- EteresDocumento2 páginasEteresJuan ManzanoAún no hay calificaciones
- Tutorial Chemsketch 2013Documento6 páginasTutorial Chemsketch 2013Paty MendozaAún no hay calificaciones
- Organic ADocumento3 páginasOrganic AMarlie OlivaresAún no hay calificaciones