También podría gustarte
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeCalificación: 4 de 5 estrellas4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe EverandThe Little Book of Hygge: Danish Secrets to Happy LivingCalificación: 3.5 de 5 estrellas3.5/5 (399)
- PFK-2 FBP-2 ReviewDocumento6 páginasPFK-2 FBP-2 Reviewapi-3710186100% (1)
- PFK 2Documento9 páginasPFK 2api-3710186Aún no hay calificaciones
- Paper002 FarmacodinámiaDocumento10 páginasPaper002 Farmacodinámiaapi-3710186100% (1)
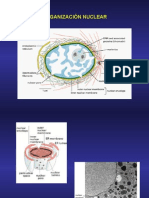
- Clase Organización Celular Biología CelularDocumento34 páginasClase Organización Celular Biología Celularapi-3710186Aún no hay calificaciones
- Paper003 FarmacodinámiaDocumento7 páginasPaper003 Farmacodinámiaapi-3710186Aún no hay calificaciones
- Clase Adhesión Biología CelularDocumento63 páginasClase Adhesión Biología Celularapi-3710186100% (1)
- Clase Genética Molecular Biología CelularDocumento45 páginasClase Genética Molecular Biología Celularapi-3710186Aún no hay calificaciones
- Clase Componentes Químicos Biología CelularDocumento46 páginasClase Componentes Químicos Biología Celularapi-3710186Aún no hay calificaciones
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryCalificación: 3.5 de 5 estrellas3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceCalificación: 4 de 5 estrellas4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)De EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Calificación: 4 de 5 estrellas4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeDe EverandShoe Dog: A Memoir by the Creator of NikeCalificación: 4.5 de 5 estrellas4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureCalificación: 4.5 de 5 estrellas4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe EverandNever Split the Difference: Negotiating As If Your Life Depended On ItCalificación: 4.5 de 5 estrellas4.5/5 (838)
- Grit: The Power of Passion and PerseveranceDe EverandGrit: The Power of Passion and PerseveranceCalificación: 4 de 5 estrellas4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaCalificación: 4.5 de 5 estrellas4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerDe EverandThe Emperor of All Maladies: A Biography of CancerCalificación: 4.5 de 5 estrellas4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealDe EverandOn Fire: The (Burning) Case for a Green New DealCalificación: 4 de 5 estrellas4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersCalificación: 4.5 de 5 estrellas4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnDe EverandTeam of Rivals: The Political Genius of Abraham LincolnCalificación: 4.5 de 5 estrellas4.5/5 (234)
- Rise of ISIS: A Threat We Can't IgnoreDe EverandRise of ISIS: A Threat We Can't IgnoreCalificación: 3.5 de 5 estrellas3.5/5 (137)
- The Unwinding: An Inner History of the New AmericaDe EverandThe Unwinding: An Inner History of the New AmericaCalificación: 4 de 5 estrellas4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyCalificación: 3.5 de 5 estrellas3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreCalificación: 4 de 5 estrellas4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Calificación: 4.5 de 5 estrellas4.5/5 (119)
- The Perks of Being a WallflowerDe EverandThe Perks of Being a WallflowerCalificación: 4.5 de 5 estrellas4.5/5 (2099)
- Her Body and Other Parties: StoriesDe EverandHer Body and Other Parties: StoriesCalificación: 4 de 5 estrellas4/5 (821)
- Math 6 Unit 8 Volume of Solids and LiquidsDocumento1 páginaMath 6 Unit 8 Volume of Solids and LiquidsRobi AkmalAún no hay calificaciones
- Steel Industry PackageDocumento9 páginasSteel Industry Packagebatung144100% (1)
- DP Level Measurement BasicsDocumento2 páginasDP Level Measurement Basicsjsrplc7952Aún no hay calificaciones
- Quantum-Mechanical Aspects of The L. Pauling's Resonance Theory.Documento4 páginasQuantum-Mechanical Aspects of The L. Pauling's Resonance Theory.Bezverkhniy VolodymyrAún no hay calificaciones
- Heat loss and fluid temperature calculation for buried insulated and non-insulated oil pipelinesDocumento34 páginasHeat loss and fluid temperature calculation for buried insulated and non-insulated oil pipelinesselisenAún no hay calificaciones
- MSDS Polyken 1027 PrimerDocumento7 páginasMSDS Polyken 1027 PrimerPungkas NisworoAún no hay calificaciones
- Vortex Quantum SeriesDocumento34 páginasVortex Quantum SeriesmiguelcAún no hay calificaciones
- Installation Details IguzziniDocumento133 páginasInstallation Details IguzziniimtiazAún no hay calificaciones
- Acid Cleaning For Nickel Alloys Pages From Handbook of Engineering Practice of Materials and Corrosion by Jung-Chul (Thomas) EunDocumento9 páginasAcid Cleaning For Nickel Alloys Pages From Handbook of Engineering Practice of Materials and Corrosion by Jung-Chul (Thomas) EunMohammad TaherAún no hay calificaciones
- 2074 1 2015 AMD2 Reff2020Documento16 páginas2074 1 2015 AMD2 Reff2020ocsspectroAún no hay calificaciones
- Product Data Booklet Fibertec Ultimate HDD Coating SystemDocumento8 páginasProduct Data Booklet Fibertec Ultimate HDD Coating Systemmkash028Aún no hay calificaciones
- CHM 1321 Assignment 1 Answers: CN H H H H HDocumento10 páginasCHM 1321 Assignment 1 Answers: CN H H H H HSara YuenAún no hay calificaciones
- Wastewater Treatment Concepts and PracticesDocumento212 páginasWastewater Treatment Concepts and Practicesfajar waskito100% (5)
- Pipe Freezing StudyDocumento8 páginasPipe Freezing StudymirekwaznyAún no hay calificaciones
- Selective Laser SinteringDocumento24 páginasSelective Laser SinteringRahul GandhiAún no hay calificaciones
- CV en - Op - Gill Giovani Awonguino oDocumento1 páginaCV en - Op - Gill Giovani Awonguino oAuguste SuelieAún no hay calificaciones
- UntitledDocumento15 páginasUntitledsahseatranAún no hay calificaciones
- Paj-7e 201906Documento2 páginasPaj-7e 201906Anum AkramAún no hay calificaciones
- Spectro RamanDocumento6 páginasSpectro RamanSampada, Astrologer and Vastu Spl. SSBAún no hay calificaciones
- Properties of Fluids: Lecture - 1Documento40 páginasProperties of Fluids: Lecture - 1Nawaz441Aún no hay calificaciones
- Construction & Material: Defects in Buildings & RemediesDocumento29 páginasConstruction & Material: Defects in Buildings & RemedieskirtikaAún no hay calificaciones
- p355n NH Nl1 Nl2 TubesDocumento4 páginasp355n NH Nl1 Nl2 Tubesado31Aún no hay calificaciones
- ANTHE 2021 (Engineering) Sample PaperDocumento17 páginasANTHE 2021 (Engineering) Sample PaperDida CowernAún no hay calificaciones
- Hygroscopic and Nonhygroscopic MaterialDocumento9 páginasHygroscopic and Nonhygroscopic Materialiresa_nuratAún no hay calificaciones
- B705 Acpt1003Documento3 páginasB705 Acpt1003taya1401Aún no hay calificaciones
- Fermi DistrbutionDocumento17 páginasFermi DistrbutionnazarAún no hay calificaciones
- B.pharm. Class NotesDocumento817 páginasB.pharm. Class NotesMukesh TiwariAún no hay calificaciones
- Heat Exchanger Mid-Term ReportDocumento18 páginasHeat Exchanger Mid-Term ReportkoanakistAún no hay calificaciones
- Epogen Safety Data Sheet 20130213 Rev 4Documento8 páginasEpogen Safety Data Sheet 20130213 Rev 4Michael Marc Andico DeleonAún no hay calificaciones
- 1 SR Star Jee Main GTM 02 - 03 01 2024 KeyDocumento14 páginas1 SR Star Jee Main GTM 02 - 03 01 2024 Keyjahnavimogarala9Aún no hay calificaciones