También podría gustarte
- Estructura y función cardiovascularDocumento19 páginasEstructura y función cardiovascularMatías OrdoñezAún no hay calificaciones
- BiomoléculasDocumento33 páginasBiomoléculasMaria Isabel Valdivia SanchezAún no hay calificaciones
- Los Ácidos NucleicosDocumento28 páginasLos Ácidos NucleicosEsther MorrásAún no hay calificaciones
- Guia ReplicacionDocumento7 páginasGuia Replicacionrosa amasAún no hay calificaciones
- Comunicación CelularDocumento43 páginasComunicación CelularDiana AguilarAún no hay calificaciones
- Taller Mitocondria 2019 IDocumento8 páginasTaller Mitocondria 2019 IJESUS ALBERTO SÁNCHEZ GARZONAún no hay calificaciones
- Funciones Del AmpcDocumento3 páginasFunciones Del AmpcRaulAún no hay calificaciones
- Cinética y Mecanismos Del Edema Celular Isosmótico Mediado Por Amoníaco y Amonio en Células de NeuroblastomaDocumento147 páginasCinética y Mecanismos Del Edema Celular Isosmótico Mediado Por Amoníaco y Amonio en Células de NeuroblastomavictorMB100% (1)
- Practico Nº 3Documento3 páginasPractico Nº 3Maria Rodriguez Olivera100% (1)
- Comunicación CelularDocumento49 páginasComunicación CelularRattus RattusAún no hay calificaciones
- Envoltura NuclearDocumento2 páginasEnvoltura NuclearKatiuska SumozaAún no hay calificaciones
- ANEXO 1. CÉLULA EUCARIOTA CorregidoDocumento2 páginasANEXO 1. CÉLULA EUCARIOTA CorregidoabAún no hay calificaciones
- Célula EucariotaDocumento8 páginasCélula EucariotaSapmcoolAún no hay calificaciones
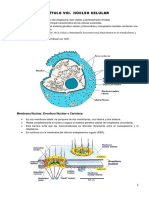
- Capítulo Viii. Núcleo CelularDocumento10 páginasCapítulo Viii. Núcleo CelularErica SaniAún no hay calificaciones
- PrÁctica 2 Célula InterfÁsica y en DivisiÓn09Documento3 páginasPrÁctica 2 Célula InterfÁsica y en DivisiÓn09esm2cm15f7031100% (2)
- Organelos de La Célula Animal y VegetalDocumento3 páginasOrganelos de La Célula Animal y Vegetalruz marina garrido De Castillo100% (1)
- Cultivos celulares: entorno y técnicasDocumento27 páginasCultivos celulares: entorno y técnicasSofía Parra ScafiezzoAún no hay calificaciones
- Citoesqueleto, ResumenDocumento13 páginasCitoesqueleto, ResumenMKsander GarcíaAún no hay calificaciones
- Glosario Biología MolecularDocumento7 páginasGlosario Biología MolecularValeria100% (1)
- Acidos Nucleicos Biologia IIDocumento31 páginasAcidos Nucleicos Biologia IIJunior Antonio Turizo100% (1)
- Tema 6. La Clasificación de Los Seres VivosDocumento29 páginasTema 6. La Clasificación de Los Seres VivosAna Vanessa Cabrera VelazquezAún no hay calificaciones
- Historia de La CélulaDocumento7 páginasHistoria de La CélulaAlender Jesus Vargas TovarAún no hay calificaciones
- Introducción a la Mitosis y MeiosisDocumento5 páginasIntroducción a la Mitosis y MeiosisSneyder Leyton Cárdenas100% (1)
- Ciclo reproductivo virus: fases fijación, eclipse, multiplicación y liberaciónDocumento5 páginasCiclo reproductivo virus: fases fijación, eclipse, multiplicación y liberaciónCristofer LopezAún no hay calificaciones
- 03fisio OcularDocumento6 páginas03fisio OcularJOELAún no hay calificaciones
- Cromosmas. Mitosis y MeiosisDocumento14 páginasCromosmas. Mitosis y MeiosisSergio Hernandez100% (1)
- Como Se Forman Las MoleculasDocumento11 páginasComo Se Forman Las MoleculasChristian ColloguazoAún no hay calificaciones
- División CelularDocumento10 páginasDivisión CelularAndrea Fernanda Calderon CuadrosAún no hay calificaciones
- CitoesqueletoDocumento50 páginasCitoesqueletoBrian Fariña100% (1)
- Acidos Nucleicos y MutacionesDocumento23 páginasAcidos Nucleicos y Mutacionesluiferalxeliu1Aún no hay calificaciones
- Partes CromosomaDocumento4 páginasPartes Cromosomalorenyis maitaAún no hay calificaciones
- Moléculas de La Adhesión CelularDocumento21 páginasMoléculas de La Adhesión CelularCarolina Romero100% (1)
- Organelos celulares eucariontesDocumento6 páginasOrganelos celulares eucariontesMauricio Palacio RojasAún no hay calificaciones
- Estructura Del ADNDocumento6 páginasEstructura Del ADNmillerlandy gutierrezAún no hay calificaciones
- Modelos de Replicacion Del ADNDocumento14 páginasModelos de Replicacion Del ADNAmbar GabrielaAún no hay calificaciones
- Mecanismos de Señalizacion de Insulina, Glucolisis, GluconeogenesisDocumento19 páginasMecanismos de Señalizacion de Insulina, Glucolisis, Gluconeogenesisxlz100% (1)
- OrganelosDocumento8 páginasOrganelosmarijo artAún no hay calificaciones
- Vitaminas y PolímerosDocumento26 páginasVitaminas y PolímerosAlvaro Rodrigo Cherres CuevaAún no hay calificaciones
- La Division CelularDocumento15 páginasLa Division CelularGustavo Andres Moreno R.100% (2)
- 1a. - Alumno - 2a. Parte - Reproducción y Herencia HumanaDocumento4 páginas1a. - Alumno - 2a. Parte - Reproducción y Herencia HumanaSandra RodriguezAún no hay calificaciones
- ADN, Genes y ProteínasDocumento8 páginasADN, Genes y ProteínasRhelbamAún no hay calificaciones
- Division CelularDocumento7 páginasDivision CelularChristian SumariaAún no hay calificaciones
- Cápsula InternaDocumento5 páginasCápsula InternaAnthony Romero100% (2)
- Estructuras y Organelos de Células EucariotasDocumento16 páginasEstructuras y Organelos de Células EucariotasJossary ButtowskiAún no hay calificaciones
- El núcleo celular: Centro de control genéticoDocumento28 páginasEl núcleo celular: Centro de control genéticoJuanCBAún no hay calificaciones
- La CelulaDocumento29 páginasLa CelulaYanira Soto MedinaAún no hay calificaciones
- Sistema de EndomembranasDocumento27 páginasSistema de EndomembranasSofía TorresAún no hay calificaciones
- Transcripción y Traduccion 01Documento24 páginasTranscripción y Traduccion 01Ricardo Medina MedicinaAún no hay calificaciones
- Biología Celular y MolecularDocumento80 páginasBiología Celular y MolecularMarylauraAún no hay calificaciones
- TALLER No 1 LA CÉLULA PROCARIOTA Y EUCARIOTADocumento2 páginasTALLER No 1 LA CÉLULA PROCARIOTA Y EUCARIOTALorena Guzman100% (1)
- Los Organelos y EstructurasDocumento3 páginasLos Organelos y EstructurasMaría Alejandra Oyanadel GuerreroAún no hay calificaciones
- Reproducción asexual y sexual en animalesDocumento6 páginasReproducción asexual y sexual en animalesMary Cruz Jurado BenavidesAún no hay calificaciones
- CICLO DE KREBS FLDocumento32 páginasCICLO DE KREBS FLGabriela CarmonaAún no hay calificaciones
- Interacciones Entre Celulas y Matriz ExtracelularDocumento10 páginasInteracciones Entre Celulas y Matriz ExtracelularIngridAún no hay calificaciones
- Señalización Celular - Matriz ExtracelularDocumento7 páginasSeñalización Celular - Matriz ExtracelularEnzo Guillermo RamirezAún no hay calificaciones
- M.A. Biologia Celular y MolecularDocumento34 páginasM.A. Biologia Celular y MolecularjoweesAún no hay calificaciones
- Capitulo 4 Membrana PlasmaticaDocumento40 páginasCapitulo 4 Membrana PlasmaticayorleAún no hay calificaciones
- Medios de cultivo vegetal: componentes esencialesDocumento25 páginasMedios de cultivo vegetal: componentes esencialesNeyser Llanos CamposAún no hay calificaciones
- Introducción A La SeñalizaciónDocumento14 páginasIntroducción A La SeñalizaciónRafael Angel Mejias IzquierdoAún no hay calificaciones
- 2.2 Procesos de Comunicación Celular.Documento33 páginas2.2 Procesos de Comunicación Celular.Jose Manuel Sanchez PachecoAún no hay calificaciones
- Antonio at As - Parasitolog A M Dica-Mediterr Neo 1998Documento310 páginasAntonio at As - Parasitolog A M Dica-Mediterr Neo 1998Gisela ReyesAún no hay calificaciones
- BulimiaDocumento2 páginasBulimiaAelita RuseelAún no hay calificaciones
- ¿Qué Es El Kéfir.Documento8 páginas¿Qué Es El Kéfir.Aelita RuseelAún no hay calificaciones
- Ultraprocesados y Mejora de La Composición de Alimentos 2021 - 2022Documento4 páginasUltraprocesados y Mejora de La Composición de Alimentos 2021 - 2022Aelita RuseelAún no hay calificaciones
- 1 FisioDocumento13 páginas1 FisioAelita RuseelAún no hay calificaciones
- Poema JDocumento1 páginaPoema JAelita RuseelAún no hay calificaciones
- EL TRABAJO VA AQUÍ: Terapias Nutricionales en La Retinopatía DiabéticaDocumento13 páginasEL TRABAJO VA AQUÍ: Terapias Nutricionales en La Retinopatía DiabéticaAelita RuseelAún no hay calificaciones
- RECOPILACIÓN PRUEBAS EXAMEN TEMAS 6,7,8y 9Documento66 páginasRECOPILACIÓN PRUEBAS EXAMEN TEMAS 6,7,8y 9Aelita RuseelAún no hay calificaciones
- INTRODUCCIÓNDocumento1 páginaINTRODUCCIÓNAelita RuseelAún no hay calificaciones
- Tema 8Documento46 páginasTema 8Aelita RuseelAún no hay calificaciones
- InstruccionesDocumento2 páginasInstruccionesAelita RuseelAún no hay calificaciones
- Tema 2. Membrana Plasmática PDFDocumento16 páginasTema 2. Membrana Plasmática PDFAelita RuseelAún no hay calificaciones
- Para La Conclusión RebecaDocumento2 páginasPara La Conclusión RebecaAelita RuseelAún no hay calificaciones
- Tema 8Documento46 páginasTema 8Aelita RuseelAún no hay calificaciones
- Tema 9Documento22 páginasTema 9Aelita RuseelAún no hay calificaciones
- Tema 7 (I) CTA. Control de Microorganismos Agentes Físicos.2019Documento30 páginasTema 7 (I) CTA. Control de Microorganismos Agentes Físicos.2019Aelita RuseelAún no hay calificaciones
- Peroxisomas PDFDocumento3 páginasPeroxisomas PDFAelita RuseelAún no hay calificaciones
- Tema 1. Introducción PDFDocumento9 páginasTema 1. Introducción PDFAelita RuseelAún no hay calificaciones
- El Ciclo Celular PDFDocumento5 páginasEl Ciclo Celular PDFAelita RuseelAún no hay calificaciones
- El Ciclo Celular PDFDocumento5 páginasEl Ciclo Celular PDFAelita RuseelAún no hay calificaciones
- BulimiaDocumento2 páginasBulimiaAelita RuseelAún no hay calificaciones
- Peroxisomas PDFDocumento3 páginasPeroxisomas PDFAelita RuseelAún no hay calificaciones
- Fundamentos de la ciencia PopperDocumento6 páginasFundamentos de la ciencia PopperAelita RuseelAún no hay calificaciones
- Wuolah Free Tema 4 Limites de La Ciencia 1Documento3 páginasWuolah Free Tema 4 Limites de La Ciencia 1Aelita RuseelAún no hay calificaciones
- Tema 10 - El Ciclo Celular 2018-2019 CASTELLANODocumento49 páginasTema 10 - El Ciclo Celular 2018-2019 CASTELLANOAelita RuseelAún no hay calificaciones
- Tema 10 - El Ciclo Celular 2018-2019 CASTELLANODocumento49 páginasTema 10 - El Ciclo Celular 2018-2019 CASTELLANOAelita RuseelAún no hay calificaciones
- COLOIDESDocumento11 páginasCOLOIDESAelita Ruseel0% (1)
- Test 5Documento10 páginasTest 5Aelita RuseelAún no hay calificaciones
- wuolah-free-LIBRO PROBLEMAS PDFDocumento256 páginaswuolah-free-LIBRO PROBLEMAS PDFAelita RuseelAún no hay calificaciones
- Guia Diagnostico y Tratamiento de La Osteoartrosis FelinaDocumento86 páginasGuia Diagnostico y Tratamiento de La Osteoartrosis FelinaSolange Valenzuela OlivaAún no hay calificaciones
- Sistema nervioso: la red que controla el cuerpoDocumento56 páginasSistema nervioso: la red que controla el cuerpoDe La Tribu NipiAún no hay calificaciones
- Neurofarmacologia Cap 1 PDFDocumento23 páginasNeurofarmacologia Cap 1 PDFFranco ToribioAún no hay calificaciones
- TIPOS DE CELULA CuadroDocumento1 páginaTIPOS DE CELULA CuadroMahly PonceAún no hay calificaciones
- Asesinato Suicida - Keith AblowDocumento900 páginasAsesinato Suicida - Keith AblowGreciaLimAún no hay calificaciones
- Sinapsis y Circuitos NeuronalesDocumento52 páginasSinapsis y Circuitos NeuronalesTery Vasquez Hassinger100% (1)
- Seperiza Ivan - Optimizando El Poder de Su MenteDocumento44 páginasSeperiza Ivan - Optimizando El Poder de Su MenteJavier RuizAún no hay calificaciones
- S3 Neuromarketing - Mas Alla Del Posicionamiento-UNFVDocumento40 páginasS3 Neuromarketing - Mas Alla Del Posicionamiento-UNFVLuis Felipe Amaut Mere100% (1)
- Anatomía InsectilDocumento140 páginasAnatomía InsectilFranklin J Urrutia Vidarte100% (1)
- Tinnitus TraducidoDocumento299 páginasTinnitus TraducidoJessy FerreiraAún no hay calificaciones
- Compendio Área Cosmovisiones, Filosofía y Sicología UsDocumento132 páginasCompendio Área Cosmovisiones, Filosofía y Sicología UsJose Luis UsnayoAún no hay calificaciones
- Biologia del comportament: Estructura y funcionamiento neuronalDocumento17 páginasBiologia del comportament: Estructura y funcionamiento neuronalCarla Carulla CapdevilaAún no hay calificaciones
- Sistema NerviosoDocumento5 páginasSistema NerviosoLaura SeoaneAún no hay calificaciones
- Mnr502 s3 Taub Others - En.esDocumento10 páginasMnr502 s3 Taub Others - En.esfelipitodenicoleAún no hay calificaciones
- Módulo 2 Estrategias de Aprendizaje Con Base en Las NeurocienciasDocumento54 páginasMódulo 2 Estrategias de Aprendizaje Con Base en Las NeurocienciasLiliana MarchenaAún no hay calificaciones
- CAP 1 Libro de Catedra.Documento9 páginasCAP 1 Libro de Catedra.Martu ContiAún no hay calificaciones
- Ipseidad y LenguajeDocumento21 páginasIpseidad y LenguajeFernando Hidalgo100% (1)
- 100000PS25 BasesBiologicasDelComportamientoDocumento5 páginas100000PS25 BasesBiologicasDelComportamientoSarela Velasquez CarrascoAún no hay calificaciones
- M1 - Desarrollo Del Lenguaje y La ComunicaciónDocumento24 páginasM1 - Desarrollo Del Lenguaje y La Comunicaciónjanet gaeteAún no hay calificaciones
- Resumen Videos Psicofisio ADocumento48 páginasResumen Videos Psicofisio AneoDavid 14Aún no hay calificaciones
- Primer Parcial de Anatomia y FisiologiaDocumento7 páginasPrimer Parcial de Anatomia y FisiologiaLu LuAún no hay calificaciones
- Todo niño puede cantarDocumento11 páginasTodo niño puede cantarCultura Comuna De Máximo PazAún no hay calificaciones
- Clip - Experimento Psicofisiologia PDFDocumento4 páginasClip - Experimento Psicofisiologia PDFJeiry CastañedaAún no hay calificaciones
- Neurociencia del aprendizaje: estructura cerebral y bases fisiológicasDocumento9 páginasNeurociencia del aprendizaje: estructura cerebral y bases fisiológicasbohane santos100% (1)
- Unidad 1 y 2Documento207 páginasUnidad 1 y 2EdwinAún no hay calificaciones
- Tipos de Células y Sus CaracteristicasDocumento6 páginasTipos de Células y Sus CaracteristicaslyamackermanAún no hay calificaciones
- Documento A4 Notas Bloc Agenda Apuntes Doodle BlancoDocumento16 páginasDocumento A4 Notas Bloc Agenda Apuntes Doodle BlancoRuth HombreAún no hay calificaciones
- SK Psicología U2 ApuntesDocumento20 páginasSK Psicología U2 ApuntesMónica PueblaAún no hay calificaciones
- Matlab PerceptronDocumento12 páginasMatlab PerceptrongomezjeffersonAún no hay calificaciones
- Realidad Virtual para Rehabilitación Física y Motora PDFDocumento165 páginasRealidad Virtual para Rehabilitación Física y Motora PDFStephannieVMGAún no hay calificaciones
- Resetea tu mente. Descubre de lo que eres capazDe EverandResetea tu mente. Descubre de lo que eres capazCalificación: 5 de 5 estrellas5/5 (195)
- Maneja tus emociones: Curso intensivo de mindfulness y regulación emocionalDe EverandManeja tus emociones: Curso intensivo de mindfulness y regulación emocionalCalificación: 4.5 de 5 estrellas4.5/5 (140)
- Cómo hacer que te pasen cosas buenas: Entiende tu cerebro, gestiona tus emociones, mejora tu vidaDe EverandCómo hacer que te pasen cosas buenas: Entiende tu cerebro, gestiona tus emociones, mejora tu vidaCalificación: 5 de 5 estrellas5/5 (1864)
- Pensamiento Crítico: Utiliza modelos mentales para desarrollar tomas de decisiones efectivas y habilidades de resolución de problemas. Supera los obstáculos cognitivos y las falacias en los sistemas para pensar con claridad en tu vida cotidiana.De EverandPensamiento Crítico: Utiliza modelos mentales para desarrollar tomas de decisiones efectivas y habilidades de resolución de problemas. Supera los obstáculos cognitivos y las falacias en los sistemas para pensar con claridad en tu vida cotidiana.Calificación: 4.5 de 5 estrellas4.5/5 (33)
- Disciplina con amor para adolescentes (Discipline With Love for Adolescents): Guía para llevarte bien con tu adolescente (A Guide for Getting Along Well With Your Adolescent)De EverandDisciplina con amor para adolescentes (Discipline With Love for Adolescents): Guía para llevarte bien con tu adolescente (A Guide for Getting Along Well With Your Adolescent)Calificación: 5 de 5 estrellas5/5 (10)
- La revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaDe EverandLa revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaCalificación: 5 de 5 estrellas5/5 (200)
- El poder del optimismo: Herramientas para vivir de forma más positivaDe EverandEl poder del optimismo: Herramientas para vivir de forma más positivaCalificación: 4.5 de 5 estrellas4.5/5 (15)
- Terapia cognitivo-conductual (TCC) y terapia dialéctico-conductual (TDC): Cómo la TCC, la TDC y la ACT pueden ayudarle a superar la ansiedad, la depresión, y los TOCSDe EverandTerapia cognitivo-conductual (TCC) y terapia dialéctico-conductual (TDC): Cómo la TCC, la TDC y la ACT pueden ayudarle a superar la ansiedad, la depresión, y los TOCSCalificación: 5 de 5 estrellas5/5 (1)
- Tu cerebro emocional: Saca partido de lo que sientes y transforma tu vidaDe EverandTu cerebro emocional: Saca partido de lo que sientes y transforma tu vidaCalificación: 5 de 5 estrellas5/5 (2)
- Psiconeuroinmunología para la práctica clínicaDe EverandPsiconeuroinmunología para la práctica clínicaCalificación: 5 de 5 estrellas5/5 (4)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Resumen de El Sutil Arte de que te Importe un Carajo, de Mark MansonDe EverandResumen de El Sutil Arte de que te Importe un Carajo, de Mark MansonCalificación: 4.5 de 5 estrellas4.5/5 (15)
- ¿Por qué mis padres no me aman?: Empezando a sanarDe Everand¿Por qué mis padres no me aman?: Empezando a sanarCalificación: 4.5 de 5 estrellas4.5/5 (33)
- Te odio - no me abandones: Comprender el trastorno límite de personalidadDe EverandTe odio - no me abandones: Comprender el trastorno límite de personalidadCalificación: 5 de 5 estrellas5/5 (3)
- Interpretación del ECG: Una Guía Práctica e Intuitiva para Aprender a Leer el ECG y Diagnosticar y Tratar ArritmiasDe EverandInterpretación del ECG: Una Guía Práctica e Intuitiva para Aprender a Leer el ECG y Diagnosticar y Tratar ArritmiasAún no hay calificaciones
- Influencia. La psicología de la persuasiónDe EverandInfluencia. La psicología de la persuasiónCalificación: 4.5 de 5 estrellas4.5/5 (14)
- No desperdicies tus emociones: Cómo lo que sientes te acerca a Dios y le da gloriaDe EverandNo desperdicies tus emociones: Cómo lo que sientes te acerca a Dios y le da gloriaCalificación: 4.5 de 5 estrellas4.5/5 (11)
- Nunca fuimos modernos: Ensayos de antropología simétricaDe EverandNunca fuimos modernos: Ensayos de antropología simétricaAún no hay calificaciones
- Disciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreDe EverandDisciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreCalificación: 5 de 5 estrellas5/5 (3)
- Cómo terminar lo que empiezas: El arte de perseverar, pasar a la acción, ejecutar los planes y tener disciplinaDe EverandCómo terminar lo que empiezas: El arte de perseverar, pasar a la acción, ejecutar los planes y tener disciplinaCalificación: 4.5 de 5 estrellas4.5/5 (6)
- Guía para superar los pensamientos atemorizantes, obsesivos o inquietantes: Libérate de los pensamientos negativos con la terapia cognitivo conductual (CBT)De EverandGuía para superar los pensamientos atemorizantes, obsesivos o inquietantes: Libérate de los pensamientos negativos con la terapia cognitivo conductual (CBT)Calificación: 4 de 5 estrellas4/5 (24)
- Cómo Desintoxicar el Cuerpo de Metales Pesados: El Secreto Poco Hablado para Sentirte Mejor, Bajar de Peso y Tener más EnergíaDe EverandCómo Desintoxicar el Cuerpo de Metales Pesados: El Secreto Poco Hablado para Sentirte Mejor, Bajar de Peso y Tener más EnergíaCalificación: 4.5 de 5 estrellas4.5/5 (2)
- Ansiedad infantil. Los trastornos explicados a los padresDe EverandAnsiedad infantil. Los trastornos explicados a los padresCalificación: 4.5 de 5 estrellas4.5/5 (25)
- Teoría polivagal práctica y terapiaDe EverandTeoría polivagal práctica y terapiaCalificación: 5 de 5 estrellas5/5 (4)