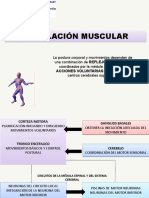
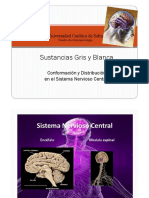
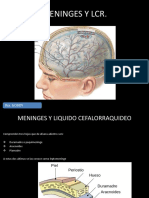
También podría gustarte
- Introducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludDe EverandIntroducción Básica a la Microcirculación de la Sangre: La Siguiente Generación en Cuidados de la SaludAún no hay calificaciones
- Músculos de La Cintura EscapularDocumento5 páginasMúsculos de La Cintura Escapularkarla daniel100% (1)
- Rehabilitación de La Artroplastia Total de Cadera - Progesión y Restricciones (7 Páginas)Documento7 páginasRehabilitación de La Artroplastia Total de Cadera - Progesión y Restricciones (7 Páginas)Walter Steve Pena AyastaAún no hay calificaciones
- Qué Es El Método de RoodDocumento13 páginasQué Es El Método de RoodalvaroAún no hay calificaciones
- Tema 15. Faradización Y Curvas (I/T) (A/T) : Lesiones de Nervios PeriféricosDocumento3 páginasTema 15. Faradización Y Curvas (I/T) (A/T) : Lesiones de Nervios PeriféricosJustine DauphinAún no hay calificaciones
- Músculos Grupo 3Documento17 páginasMúsculos Grupo 3K Gallegos Sedeño PalomaAún no hay calificaciones
- Evidencia de Las Ondas de ChoqueDocumento7 páginasEvidencia de Las Ondas de ChoqueamexatoAún no hay calificaciones
- Control Motor y PlanificaciónDocumento16 páginasControl Motor y Planificaciónyamile_love_167450% (2)
- Cuadro de Los Tipos de ColagenoDocumento6 páginasCuadro de Los Tipos de ColagenoVictor hugo AbadiaAún no hay calificaciones
- Rcqconsolidación Del Hueso Mark DuttonDocumento61 páginasRcqconsolidación Del Hueso Mark DuttonAldo NavarreteAún no hay calificaciones
- Via Piramidal y ReflejosDocumento36 páginasVia Piramidal y ReflejosEdwin Calle100% (2)
- Inestabilidad Cronica TobilloDocumento12 páginasInestabilidad Cronica TobilloSebastián VilchesAún no hay calificaciones
- Musculo Esquelético y Su ContracciónDocumento33 páginasMusculo Esquelético y Su ContracciónPaolaAún no hay calificaciones
- APUNTES EmbriologiaDocumento5 páginasAPUNTES Embriologiagabriel ChuquimiaAún no hay calificaciones
- SO.001. Exploración de Hombro Doloroso. Revista Jano, 2008.Documento5 páginasSO.001. Exploración de Hombro Doloroso. Revista Jano, 2008.Alejandro Guzman0% (1)
- Modelos InterpersonalesDocumento14 páginasModelos InterpersonalesAnonymous oAGumZoAún no hay calificaciones
- Lumbalgia MecánicaDocumento3 páginasLumbalgia MecánicaJorge GomezAún no hay calificaciones
- Cuestionario de MúsculoDocumento10 páginasCuestionario de MúsculoLa Chica de los GooglesAún no hay calificaciones
- Region Tibial PosteriorDocumento18 páginasRegion Tibial PosteriorPaolaLucioAún no hay calificaciones
- Nervio GlosofaringeoDocumento8 páginasNervio GlosofaringeoJuan Alcides Alvarez CabreraAún no hay calificaciones
- Engrama NeuronalDocumento25 páginasEngrama NeuronalDiego Campo100% (4)
- 2 - Unidad 2 y 3 ORTESIS DE TRONCODocumento11 páginas2 - Unidad 2 y 3 ORTESIS DE TRONCOAlex GarciaAún no hay calificaciones
- Marcha KlappDocumento21 páginasMarcha KlappPipe Cisternas IlabacaAún no hay calificaciones
- Regulación Muscular1Documento19 páginasRegulación Muscular1Lourdes PerdomoAún no hay calificaciones
- RODILLADocumento24 páginasRODILLAGastón ColiñirAún no hay calificaciones
- Imprimir Guía Movimientos Articulares PlanimetríaDocumento16 páginasImprimir Guía Movimientos Articulares PlanimetríaG. Lizeth Avila MonroyAún no hay calificaciones
- Corrientes GalvanicasDocumento19 páginasCorrientes GalvanicasAilin VinuezaAún no hay calificaciones
- Tonos musculares y sus implicaciones en el desarrollo motorDocumento33 páginasTonos musculares y sus implicaciones en el desarrollo motorRaúl Nolasco VelazquezAún no hay calificaciones
- Manual NeuroDocumento17 páginasManual NeuroRaul RLAún no hay calificaciones
- Músculos Del Compartimiento AnteriorDocumento31 páginasMúsculos Del Compartimiento AnteriorCristian Andres ZafraAún no hay calificaciones
- Fractura de CollesDocumento11 páginasFractura de ColleswileAún no hay calificaciones
- OsteomuscularDocumento53 páginasOsteomuscularLaura Juliana Quintero WiderkehrAún no hay calificaciones
- Rehabilitación en Enfermedades Degenerativas CerebelosasDocumento13 páginasRehabilitación en Enfermedades Degenerativas Cerebelosasjsanchez59Aún no hay calificaciones
- Exploracion de La Marcha Bueno SanchezDocumento7 páginasExploracion de La Marcha Bueno SanchezAnonymous ACw3SGPWa100% (1)
- Actividad Eléctrica Del Cerebro, Estados Del SueñoDocumento10 páginasActividad Eléctrica Del Cerebro, Estados Del SueñoAlex MorenoAún no hay calificaciones
- Receptores y efectoresDocumento32 páginasReceptores y efectoresYENIS MEJIA PAún no hay calificaciones
- Lesiones MuscularesDocumento8 páginasLesiones MuscularesMyri GuerreroAún no hay calificaciones
- Anatomía y biomecánica de los dedos de la manoDocumento63 páginasAnatomía y biomecánica de los dedos de la manoAnonymous btZNNQGrAún no hay calificaciones
- Marchas con bastones y muletas: tipos 4, 3 y 2 puntosDocumento3 páginasMarchas con bastones y muletas: tipos 4, 3 y 2 puntosVerónica Rivera EspinozaAún no hay calificaciones
- Fractura Monteggia y GaleazziDocumento4 páginasFractura Monteggia y GaleazziKaty Marlen GongoraAún no hay calificaciones
- Retención de líquidos y fases reparación tisularDocumento4 páginasRetención de líquidos y fases reparación tisularValeria HernándezAún no hay calificaciones
- Musculos Del Esqueleto Apendicular Tema VDocumento13 páginasMusculos Del Esqueleto Apendicular Tema VMilagro MonsalveAún no hay calificaciones
- Artritis ReumatoideDocumento32 páginasArtritis ReumatoideRobert GonzalezAún no hay calificaciones
- Plexo LumbarDocumento2 páginasPlexo LumbarAllan GarciaAún no hay calificaciones
- Explorando PDFDocumento6 páginasExplorando PDFRudy RamosAún no hay calificaciones
- Plexo Braquial y LumbosacroDocumento4 páginasPlexo Braquial y LumbosacroDiana LeccaAún no hay calificaciones
- Arbol BronquialDocumento1 páginaArbol BronquialLester AguilarAún no hay calificaciones
- Tractos y VíasDocumento14 páginasTractos y VíasFabiana CastañedaAún no hay calificaciones
- ARQUICEREBELODocumento5 páginasARQUICEREBELOKidnez Reyes AltamiranoAún no hay calificaciones
- Categorias, Test y MedidasDocumento5 páginasCategorias, Test y MedidasPOLARツ100% (1)
- MeniscopatiaDocumento7 páginasMeniscopatiaEnrique GarridoAún no hay calificaciones
- Cap 48 GuytonDocumento15 páginasCap 48 GuytonKaren Andrea StAún no hay calificaciones
- Goniometria de La Columna CervicalDocumento3 páginasGoniometria de La Columna CervicalPazita Fuentes JaraAún no hay calificaciones
- Kinefilaxia - Fisiología Muscular y Fuerza PDFDocumento79 páginasKinefilaxia - Fisiología Muscular y Fuerza PDFNina VillarroelAún no hay calificaciones
- Sistema Piramidal y ExtrapiramidalDocumento2 páginasSistema Piramidal y ExtrapiramidalDinaOrdoñezAún no hay calificaciones
- 06 Exploracion ColumnaDocumento27 páginas06 Exploracion ColumnaJose Luis Tabueña OrtegaAún no hay calificaciones
- Exploracion Mano y MuecaDocumento51 páginasExploracion Mano y MuecaGina DurandAún no hay calificaciones
- Tono Muscular-1Documento82 páginasTono Muscular-1Rivera Gómez América LucíaAún no hay calificaciones
- Sistema Inhibidor de La Celula de RenshawDocumento8 páginasSistema Inhibidor de La Celula de RenshawMonte Sann MarielaAún no hay calificaciones
- Qué son los reflejos muscularesDocumento8 páginasQué son los reflejos muscularesMartin reyesAún no hay calificaciones
- Circulaciones Especiales PDFDocumento52 páginasCirculaciones Especiales PDFmeli murAún no hay calificaciones
- Microcirculación: Generalidades y FuncionesDocumento32 páginasMicrocirculación: Generalidades y Funcionesmeli mur100% (1)
- Guia Presion Arterial PDFDocumento3 páginasGuia Presion Arterial PDFmeli murAún no hay calificaciones
- Regulacion Cardiovascular PDFDocumento40 páginasRegulacion Cardiovascular PDFmeli murAún no hay calificaciones
- Postura 05 de JulioDocumento3 páginasPostura 05 de Juliomeli murAún no hay calificaciones
- Circulaciones Especiales PDFDocumento52 páginasCirculaciones Especiales PDFmeli mur100% (1)
- Guia EKGDocumento1 páginaGuia EKGmeli murAún no hay calificaciones
- Ciclo cardíaco, dinámica y contractilidadDocumento59 páginasCiclo cardíaco, dinámica y contractilidadmeli murAún no hay calificaciones
- Regulacion Cardiovascular PDFDocumento40 páginasRegulacion Cardiovascular PDFmeli murAún no hay calificaciones
- Electrofisiologia CardiacaDocumento59 páginasElectrofisiologia Cardiacameli murAún no hay calificaciones
- Mecanica Cardiaca PDFDocumento59 páginasMecanica Cardiaca PDFmeli murAún no hay calificaciones
- Sistema Nervioso AutónomoDocumento13 páginasSistema Nervioso Autónomomeli murAún no hay calificaciones
- Transcripción Fisiología CelularDocumento12 páginasTranscripción Fisiología Celularmeli murAún no hay calificaciones
- Movimiento Voluntario 05 JulioDocumento4 páginasMovimiento Voluntario 05 Juliomeli murAún no hay calificaciones
- Postura y Locomoción BlancaDocumento42 páginasPostura y Locomoción Blancameli murAún no hay calificaciones
- Sistema Nervioso AutónomoDocumento13 páginasSistema Nervioso Autónomomeli murAún no hay calificaciones
- Regulacion Cortical Del Movimiento VoluntarioDocumento6 páginasRegulacion Cortical Del Movimiento Voluntariomeli murAún no hay calificaciones
- Sistema Nerviosos Autonomo PDFDocumento37 páginasSistema Nerviosos Autonomo PDFmeli murAún no hay calificaciones
- Reflejos Posturales - Movimiento VoluntarioDocumento10 páginasReflejos Posturales - Movimiento Voluntariomeli murAún no hay calificaciones
- SISTEMA NERVIOSO CentralDocumento24 páginasSISTEMA NERVIOSO Centralbill nelson rodriguez ancajimaAún no hay calificaciones
- BIOLOGÍADocumento7 páginasBIOLOGÍAMiguel OlivaresAún no hay calificaciones
- Los Lóbulos Del CerebroDocumento10 páginasLos Lóbulos Del CerebroBladiss AvalosAún no hay calificaciones
- Médula EspinalDocumento5 páginasMédula EspinalYohany Montenegro MuñozAún no hay calificaciones
- Diferentes Iconos en La Barra de Herramientas y Su FunciónDocumento2 páginasDiferentes Iconos en La Barra de Herramientas y Su FunciónjoseAún no hay calificaciones
- Medula Espinal SesionDocumento29 páginasMedula Espinal SesionArturo Gomez CanoAún no hay calificaciones
- Sistema Nervioso Periférico forma y funciónDocumento5 páginasSistema Nervioso Periférico forma y funciónwilybarrera0% (1)
- Plexo braquial: Estructura nerviosa del miembro superiorDocumento4 páginasPlexo braquial: Estructura nerviosa del miembro superiorStalinChacónAún no hay calificaciones
- Vias OpticasDocumento1 páginaVias OpticasKeren Anaya CamargoAún no hay calificaciones
- PROTUBERANCIA O PUENTE (Configuracion Interna 2° Parte)Documento20 páginasPROTUBERANCIA O PUENTE (Configuracion Interna 2° Parte)Sor EchegarayAún no hay calificaciones
- Fisiología Del Sistema Nervioso CentralDocumento1 páginaFisiología Del Sistema Nervioso CentralGiannina PradoAún no hay calificaciones
- Preguntas Caso Clinico - s2 - Siringomielia PostraumaticaDocumento2 páginasPreguntas Caso Clinico - s2 - Siringomielia Postraumaticamaria inesAún no hay calificaciones
- Tarea 2 de Anatomia y Fisiologia Del Sistema NerviosoDocumento9 páginasTarea 2 de Anatomia y Fisiologia Del Sistema NerviosoAime BélangerAún no hay calificaciones
- Sistema Nervioso CentralDocumento4 páginasSistema Nervioso CentralADRIANA PAN100% (1)
- Articulo Corteza CerebralDocumento3 páginasArticulo Corteza CerebralAntonella MarcaAún no hay calificaciones
- Puente de VarolioDocumento2 páginasPuente de VarolioAdela SofíaAún no hay calificaciones
- Pares Craneales Del 7 Al 12Documento31 páginasPares Craneales Del 7 Al 12Leslie RodasAún no hay calificaciones
- Folleto de Morfo 5Documento2 páginasFolleto de Morfo 5Rosa SanmiguelAún no hay calificaciones
- Psicología General I Tema IIDocumento4 páginasPsicología General I Tema IIDavid AriasAún no hay calificaciones
- El EncefaloDocumento10 páginasEl EncefaloYOMAIRA MENA AGUALIMPIAAún no hay calificaciones
- Funciones cerebrales examen tópico neurocienciaDocumento11 páginasFunciones cerebrales examen tópico neurocienciaELIZABETH PARDOAún no hay calificaciones
- Diencefalo 1Documento122 páginasDiencefalo 1Luis Francisco Chávez VázquezAún no hay calificaciones
- Nervio olfatorioDocumento2 páginasNervio olfatorioBrandon ZúñigaAún no hay calificaciones
- NeuroDocumento70 páginasNeuroMafe MafeAún no hay calificaciones
- Nucleos Celulares Suastancia GrisDocumento4 páginasNucleos Celulares Suastancia GrisWilliam Erick Avalos MendozaAún no hay calificaciones
- 6 Componentes Del Sistema Musculo EsqueléticoDocumento22 páginas6 Componentes Del Sistema Musculo EsqueléticoDr. Jorge Alam100% (10)
- Estructura y distribución de la sustancia gris y blanca en el encéfaloDocumento11 páginasEstructura y distribución de la sustancia gris y blanca en el encéfaloCristian Nicolás HerreraAún no hay calificaciones
- MeningesDocumento19 páginasMeningesPaolo Jesus Cotrina Robles100% (1)
- HoyDocumento2905 páginasHoyLuis VelascoAún no hay calificaciones
- Pares CranealesDocumento15 páginasPares CranealesBriset SeminoAún no hay calificaciones
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanDe EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanCalificación: 4.5 de 5 estrellas4.5/5 (63)
- Cómo hacer que te pasen cosas buenas: Entiende tu cerebro, gestiona tus emociones, mejora tu vidaDe EverandCómo hacer que te pasen cosas buenas: Entiende tu cerebro, gestiona tus emociones, mejora tu vidaCalificación: 5 de 5 estrellas5/5 (1864)
- La revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaDe EverandLa revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaCalificación: 5 de 5 estrellas5/5 (200)
- El lenguaje del cuerpo: Una guía para conocer los sentimientos y las emociones de quienes nos rodeanDe EverandEl lenguaje del cuerpo: Una guía para conocer los sentimientos y las emociones de quienes nos rodeanCalificación: 4 de 5 estrellas4/5 (49)
- Tu cerebro emocional: Saca partido de lo que sientes y transforma tu vidaDe EverandTu cerebro emocional: Saca partido de lo que sientes y transforma tu vidaCalificación: 5 de 5 estrellas5/5 (2)
- El poder del optimismo: Herramientas para vivir de forma más positivaDe EverandEl poder del optimismo: Herramientas para vivir de forma más positivaCalificación: 4.5 de 5 estrellas4.5/5 (15)
- Resetea tu mente. Descubre de lo que eres capazDe EverandResetea tu mente. Descubre de lo que eres capazCalificación: 5 de 5 estrellas5/5 (195)
- Terapia cognitivo-conductual (TCC) y terapia dialéctico-conductual (TDC): Cómo la TCC, la TDC y la ACT pueden ayudarle a superar la ansiedad, la depresión, y los TOCSDe EverandTerapia cognitivo-conductual (TCC) y terapia dialéctico-conductual (TDC): Cómo la TCC, la TDC y la ACT pueden ayudarle a superar la ansiedad, la depresión, y los TOCSCalificación: 5 de 5 estrellas5/5 (1)
- Pensamiento Crítico: Utiliza modelos mentales para desarrollar tomas de decisiones efectivas y habilidades de resolución de problemas. Supera los obstáculos cognitivos y las falacias en los sistemas para pensar con claridad en tu vida cotidiana.De EverandPensamiento Crítico: Utiliza modelos mentales para desarrollar tomas de decisiones efectivas y habilidades de resolución de problemas. Supera los obstáculos cognitivos y las falacias en los sistemas para pensar con claridad en tu vida cotidiana.Calificación: 4.5 de 5 estrellas4.5/5 (33)
- Nunca fuimos modernos: Ensayos de antropología simétricaDe EverandNunca fuimos modernos: Ensayos de antropología simétricaAún no hay calificaciones
- Disciplina con amor para adolescentes (Discipline With Love for Adolescents): Guía para llevarte bien con tu adolescente (A Guide for Getting Along Well With Your Adolescent)De EverandDisciplina con amor para adolescentes (Discipline With Love for Adolescents): Guía para llevarte bien con tu adolescente (A Guide for Getting Along Well With Your Adolescent)Calificación: 5 de 5 estrellas5/5 (10)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoDe EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoCalificación: 5 de 5 estrellas5/5 (2)
- Influencia. La psicología de la persuasiónDe EverandInfluencia. La psicología de la persuasiónCalificación: 4.5 de 5 estrellas4.5/5 (14)
- Cómo terminar lo que empiezas: El arte de perseverar, pasar a la acción, ejecutar los planes y tener disciplinaDe EverandCómo terminar lo que empiezas: El arte de perseverar, pasar a la acción, ejecutar los planes y tener disciplinaCalificación: 4.5 de 5 estrellas4.5/5 (6)
- Te odio - no me abandones: Comprender el trastorno límite de personalidadDe EverandTe odio - no me abandones: Comprender el trastorno límite de personalidadCalificación: 5 de 5 estrellas5/5 (3)
- Ansiedad infantil. Los trastornos explicados a los padresDe EverandAnsiedad infantil. Los trastornos explicados a los padresCalificación: 4.5 de 5 estrellas4.5/5 (25)
- Maneja tus emociones: Curso intensivo de mindfulness y regulación emocionalDe EverandManeja tus emociones: Curso intensivo de mindfulness y regulación emocionalCalificación: 4.5 de 5 estrellas4.5/5 (140)
- Psiconeuroinmunología para la práctica clínicaDe EverandPsiconeuroinmunología para la práctica clínicaCalificación: 5 de 5 estrellas5/5 (4)
- Interpretación del ECG: Una Guía Práctica e Intuitiva para Aprender a Leer el ECG y Diagnosticar y Tratar ArritmiasDe EverandInterpretación del ECG: Una Guía Práctica e Intuitiva para Aprender a Leer el ECG y Diagnosticar y Tratar ArritmiasAún no hay calificaciones
- Resumen de El Sutil Arte de que te Importe un Carajo, de Mark MansonDe EverandResumen de El Sutil Arte de que te Importe un Carajo, de Mark MansonCalificación: 4.5 de 5 estrellas4.5/5 (15)
- ¿Por qué mis padres no me aman?: Empezando a sanarDe Everand¿Por qué mis padres no me aman?: Empezando a sanarCalificación: 4.5 de 5 estrellas4.5/5 (33)
- Disciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreDe EverandDisciplina Mental: Técnicas infalibles para lograr todo lo que te propones y eliminar la pereza y la procrastinación de tu vida para siempreCalificación: 5 de 5 estrellas5/5 (3)
- Cómo Conversar Con Cualquier Persona: Mejora tus habilidades sociales, desarrolla tu carisma, domina las conversaciones triviales y conviértete en una persona sociable para hacer verdaderos amigos y construir relaciones significativas.De EverandCómo Conversar Con Cualquier Persona: Mejora tus habilidades sociales, desarrolla tu carisma, domina las conversaciones triviales y conviértete en una persona sociable para hacer verdaderos amigos y construir relaciones significativas.Calificación: 5 de 5 estrellas5/5 (54)
- Diagnóstico y tratamiento del canino retenido: Aspectos clavesDe EverandDiagnóstico y tratamiento del canino retenido: Aspectos clavesCalificación: 3.5 de 5 estrellas3.5/5 (9)
- La invasión de la pseudociencia: Claves para orientarse en un mundo en donde casi todo es posibleDe EverandLa invasión de la pseudociencia: Claves para orientarse en un mundo en donde casi todo es posibleCalificación: 4.5 de 5 estrellas4.5/5 (15)
- Inteligencia artificial: Lo que usted necesita saber sobre el aprendizaje automático, robótica, aprendizaje profundo, Internet de las cosas, redes neuronales, y nuestro futuroDe EverandInteligencia artificial: Lo que usted necesita saber sobre el aprendizaje automático, robótica, aprendizaje profundo, Internet de las cosas, redes neuronales, y nuestro futuroCalificación: 4 de 5 estrellas4/5 (1)