También podría gustarte
- Une-En Iso 4628-1 2004Documento10 páginasUne-En Iso 4628-1 2004Carmen Rodriguez PoloAún no hay calificaciones
- Participación de La Familia en La Educación BásicaDocumento8 páginasParticipación de La Familia en La Educación BásicaBills Arthur Wingrove LynchAún no hay calificaciones
- Introduccion A La Psicologia Social - Marin Martinez PDFDocumento221 páginasIntroduccion A La Psicologia Social - Marin Martinez PDFchanelbrito73% (11)
- TERAPIAS BREVES REFLEXIVASDocumento4 páginasTERAPIAS BREVES REFLEXIVASTarsila Cicconetti100% (1)
- Web TriageObstetricoCMDocumento70 páginasWeb TriageObstetricoCMluis guerraAún no hay calificaciones
- Piaget: Esquemas cognitivos, asimilación y acomodaciónDocumento5 páginasPiaget: Esquemas cognitivos, asimilación y acomodaciónJennifer Gordon BarredoAún no hay calificaciones
- Una Reflexion Sobre El Metasentido de La Praxis Cientifica PDFDocumento281 páginasUna Reflexion Sobre El Metasentido de La Praxis Cientifica PDFCristian jimènezAún no hay calificaciones
- Quiroz, Historia de La Psic Social PDFDocumento29 páginasQuiroz, Historia de La Psic Social PDFPatricio Estay ConstancioAún no hay calificaciones
- PsicologiaDocumento33 páginasPsicologiaBrayan Valladolid VenturaAún no hay calificaciones
- 2109 8220 1 PBDocumento15 páginas2109 8220 1 PBKelly OrozcoAún no hay calificaciones
- Revista 2Documento25 páginasRevista 2nancyAún no hay calificaciones
- Espacios en Blanco. Revista de Educación 1515-9485: IssnDocumento39 páginasEspacios en Blanco. Revista de Educación 1515-9485: IssnAndresSalasAún no hay calificaciones
- LL Lo Que Debes Saber Sobre La Psicología Comunitaria en EEUUDocumento18 páginasLL Lo Que Debes Saber Sobre La Psicología Comunitaria en EEUUYhone MoralesAún no hay calificaciones
- Aportaciones Desde La Antropologia A La CulturaDocumento21 páginasAportaciones Desde La Antropologia A La CulturaJeison Martinez DiazAún no hay calificaciones
- Antropologia y Spicolo PDFDocumento13 páginasAntropologia y Spicolo PDFNIDYA BELLOAún no hay calificaciones
- Algunos Alcances Sobre Desarrollo Consolidación Cientifica La Antropologáa PsicológzcaDocumento11 páginasAlgunos Alcances Sobre Desarrollo Consolidación Cientifica La Antropologáa PsicológzcaJamith Eduardo Osorio ManjarrezAún no hay calificaciones
- 29200907Documento20 páginas29200907Yhone MoralesAún no hay calificaciones
- Consideraciones Educativas PerspectivaDocumento14 páginasConsideraciones Educativas PerspectivaEduardo PerezAún no hay calificaciones
- Historia Psico SocialDocumento44 páginasHistoria Psico SocialLourdes MendezAún no hay calificaciones
- Fundamentos da teoria bioecológica de BronfenbrennerDocumento11 páginasFundamentos da teoria bioecológica de BronfenbrennerUkyo TachibanaAún no hay calificaciones
- 2170 8452 1 PBDocumento9 páginas2170 8452 1 PBJose Eduardo Peralta BernalAún no hay calificaciones
- 29201508Documento20 páginas29201508Yhone MoralesAún no hay calificaciones
- 29200948Documento6 páginas29200948Yhone MoralesAún no hay calificaciones
- Apuntes Sobre Una Historia Del Paradigma Dominante de La Psicología SocialDocumento10 páginasApuntes Sobre Una Historia Del Paradigma Dominante de La Psicología SocialYhone MoralesAún no hay calificaciones
- 407-Texto Del Artículo-849-1-10-20140304Documento10 páginas407-Texto Del Artículo-849-1-10-20140304Anonymous GHwLfeJ2Aún no hay calificaciones
- Aprendizaje Profundo y SuperficialDocumento2 páginasAprendizaje Profundo y SuperficialYhone Morales100% (1)
- Nefrostomia PercutaneaDocumento1 páginaNefrostomia PercutaneaYhone MoralesAún no hay calificaciones
- ConstructivismoDocumento18 páginasConstructivismoRodrigo LeónAún no hay calificaciones
- ConstructivismoDocumento18 páginasConstructivismoLaura BravoAún no hay calificaciones
- Malla PSICOLOGÍA 2018-10-17 PDFDocumento1 páginaMalla PSICOLOGÍA 2018-10-17 PDFYhone MoralesAún no hay calificaciones
- Ejemplos Referencias Bibliográficas - NormasAPADocumento3 páginasEjemplos Referencias Bibliográficas - NormasAPADemian Dennis Tipo MamaniAún no hay calificaciones
- Unidad 2 Paso 3 Fundamentacion y Diseño de Un InstrumentoDocumento37 páginasUnidad 2 Paso 3 Fundamentacion y Diseño de Un InstrumentoYhone Morales100% (2)
- Unidad 2 Paso 3 Fundamentacion y Diseño de Un InstrumentoDocumento37 páginasUnidad 2 Paso 3 Fundamentacion y Diseño de Un InstrumentoYhone Morales100% (2)
- Lectura Semana 2la Transformación de La MariposaDocumento3 páginasLectura Semana 2la Transformación de La MariposaSilvia RodríguezAún no hay calificaciones
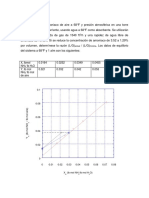
- Absorción de amoniaco en torre empacada usando agua como absorbenteDocumento6 páginasAbsorción de amoniaco en torre empacada usando agua como absorbenteNazaret Nazario RomualdoAún no hay calificaciones
- Claves de Examen Español 2Documento3 páginasClaves de Examen Español 2Lizeth AréchigaAún no hay calificaciones
- Alejandro Ramos - Antropología TeológicaDocumento23 páginasAlejandro Ramos - Antropología TeológicaRafael FabiánAún no hay calificaciones
- Creacion de PersonajeDocumento8 páginasCreacion de Personajetgv6qp7bjvAún no hay calificaciones
- Ejemplos de CondicionamientoDocumento4 páginasEjemplos de Condicionamientojezabel castilloAún no hay calificaciones
- 1 Atomo 2020Documento1 página1 Atomo 2020Micaela GimenezAún no hay calificaciones
- Coeficiente de DifusiónDocumento3 páginasCoeficiente de DifusiónAlison Yineth Montero CollazosAún no hay calificaciones
- FLEXIÓN SIMPLE NORMAL Y OBLÍCUA EN VIGAS Y ESTRUCTURASDocumento46 páginasFLEXIÓN SIMPLE NORMAL Y OBLÍCUA EN VIGAS Y ESTRUCTURASphiliphAún no hay calificaciones
- Biofisica 2021Documento51 páginasBiofisica 2021Mel LantesAún no hay calificaciones
- Tecnico de LabDocumento4 páginasTecnico de LabpanaderaAún no hay calificaciones
- Plantilla - Entrega 1-Estrategias GerencialesDocumento7 páginasPlantilla - Entrega 1-Estrategias GerencialesWILSON FERNANDO LAGOS MORERAAún no hay calificaciones
- Atimanual Del Mal Historiador ReseñaDocumento3 páginasAtimanual Del Mal Historiador ReseñaCristian TrejosAún no hay calificaciones
- Desafío Kunan 2020 4Documento2 páginasDesafío Kunan 2020 4Styp Zuñiga GuillenAún no hay calificaciones
- Red de Desague Alfonso UgarteDocumento5 páginasRed de Desague Alfonso UgarteKlisbert Ayrton Beltran LauraAún no hay calificaciones
- Planeación estratégica: horizonte y tipos enDocumento5 páginasPlaneación estratégica: horizonte y tipos enPatricia Rios100% (1)
- 1.2 ConveccionDocumento7 páginas1.2 ConveccionLuis Ramon Ramon CanelaAún no hay calificaciones
- Desarrollo de La Tecnica de Pulso Galvanostatico PDocumento8 páginasDesarrollo de La Tecnica de Pulso Galvanostatico PAndrea PerezAún no hay calificaciones
- Tipos de MuestreoDocumento29 páginasTipos de MuestreoevelynAún no hay calificaciones
- Prueba Escrita EticaDocumento2 páginasPrueba Escrita Eticamarisol lopezAún no hay calificaciones
- Act 4Documento18 páginasAct 4Gom CarletteAún no hay calificaciones
- TPA 01 F1 Aspectos - Ambientales D04Documento2 páginasTPA 01 F1 Aspectos - Ambientales D04Luis Felipe JimenezAún no hay calificaciones
- Clase 2 - Introducción A La Gerencia de Proyectos Bajo La Metodología PMI®Documento23 páginasClase 2 - Introducción A La Gerencia de Proyectos Bajo La Metodología PMI®Aleja VillalobosAún no hay calificaciones
- HumedadDocumento18 páginasHumedadPatricia CastroAún no hay calificaciones
- Determinación de Ácido AscórbicoDocumento3 páginasDeterminación de Ácido AscórbicoJorge RomoAún no hay calificaciones
- Proyecto 6 Semana 31 de SextoDocumento4 páginasProyecto 6 Semana 31 de SextoCristian Guayracaja100% (1)