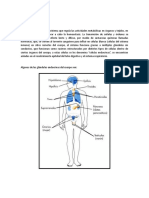
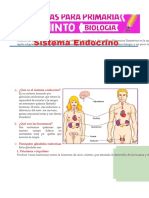
También podría gustarte
- 16 Modulos Especificos Del EXANI II PsicologiaDocumento9 páginas16 Modulos Especificos Del EXANI II PsicologiaJoel F CstlAún no hay calificaciones
- Glándulas AnexasDocumento8 páginasGlándulas AnexasMartha DominguezAún no hay calificaciones
- Cáncer de Tiroides JCDocumento21 páginasCáncer de Tiroides JCJuan Carlos MedinaAún no hay calificaciones
- Anatomía y Fisiología Renal PDFDocumento57 páginasAnatomía y Fisiología Renal PDFMinastauriel Alassëa100% (7)
- Yoga para la Mujer: Equilibrio físico y mental a través del ciclo de vidaDocumento16 páginasYoga para la Mujer: Equilibrio físico y mental a través del ciclo de vidaAnabella JimenezAún no hay calificaciones
- CO 16 Glandulas EndocrinasDocumento82 páginasCO 16 Glandulas EndocrinasJatnielAún no hay calificaciones
- Hipoglicemia NeonatalDocumento26 páginasHipoglicemia Neonatalper7000Aún no hay calificaciones
- Contenido 10 Sistema EndocrinoDocumento10 páginasContenido 10 Sistema EndocrinoEricka Paucar Troncos100% (1)
- Tejidos AnimalesDocumento21 páginasTejidos AnimalesSara GarcíaAún no hay calificaciones
- Tarea Unidad III y IV Sexualidad HumanaDocumento22 páginasTarea Unidad III y IV Sexualidad HumanaNicole AbreuAún no hay calificaciones
- Glosario de Las GlandulasDocumento5 páginasGlosario de Las GlandulasFrancisco TorresAún no hay calificaciones
- Sistema EndocrinoDocumento22 páginasSistema EndocrinoMAXIMILIANO ENRIQUE ACRAAún no hay calificaciones
- Sistema Endocrino (Resumen Ross)Documento6 páginasSistema Endocrino (Resumen Ross)Miguel IA100% (5)
- Sistema Endocrino Universidad Mariano Gálvez GuatemalaDocumento12 páginasSistema Endocrino Universidad Mariano Gálvez GuatemalaLuis Felipe MazariegosAún no hay calificaciones
- Histologia Sistema EndocrinoDocumento5 páginasHistologia Sistema EndocrinochevelosoAún no hay calificaciones
- Presentación 4 - Glándulas EndocrinasDocumento15 páginasPresentación 4 - Glándulas EndocrinasMarlon Yasir Moreno GomezAún no hay calificaciones
- Sistema Endocrino (1) (1) ZXDocumento75 páginasSistema Endocrino (1) (1) ZXLuck ScordAún no hay calificaciones
- Sistema EndocrinoDocumento8 páginasSistema EndocrinoMatiasAún no hay calificaciones
- Sistema EndocrinoDocumento11 páginasSistema EndocrinoLisdee BelloAún no hay calificaciones
- Org. Endocrinos 21 CDocumento22 páginasOrg. Endocrinos 21 CBelinda Sarai Mancilla RivasAún no hay calificaciones
- HistoDocumento14 páginasHistoisabella jimenez cadavidAún no hay calificaciones
- Lugares de producción de las principales hormonas endocrinasDocumento5 páginasLugares de producción de las principales hormonas endocrinasleonardoperezAún no hay calificaciones
- Características Bioquímicas y Morfológicas Del Sistema Endocrino PDFDocumento15 páginasCaracterísticas Bioquímicas y Morfológicas Del Sistema Endocrino PDFDanielaAún no hay calificaciones
- Cuestionarios Endocrino, Sanguineo y LinfaticoDocumento23 páginasCuestionarios Endocrino, Sanguineo y LinfaticoReymundo SeguraAún no hay calificaciones
- Unidad Educativa Dionicio Morales ChoqueDocumento8 páginasUnidad Educativa Dionicio Morales ChoqueSERVITEC CELAún no hay calificaciones
- Sistema Endocrino - TrabajoDocumento11 páginasSistema Endocrino - TrabajoYorley barbosaAún no hay calificaciones
- Endocrino - HistologíaDocumento59 páginasEndocrino - HistologíaRodrigo Cañete100% (1)
- Facultad de Ciencias de La Salud Escuela Profesional de EnfermeríaDocumento17 páginasFacultad de Ciencias de La Salud Escuela Profesional de EnfermeríaANLLY NICOL SILVA SULLONAún no hay calificaciones
- Lab5 EndoDocumento18 páginasLab5 EndoJoshua MenesesAún no hay calificaciones
- Sistema endocrinoDocumento7 páginasSistema endocrinoKristel GarciaAún no hay calificaciones
- Sistema EndocrinoDocumento37 páginasSistema EndocrinoJoselin America Pérez PolancoAún no hay calificaciones
- Histología Del Sistema EndocrinoDocumento33 páginasHistología Del Sistema EndocrinoCarlosAún no hay calificaciones
- Resumen EndocrinoDocumento9 páginasResumen EndocrinoAgustin SarAún no hay calificaciones
- European Day of The Prevention of The Cardiovascular Risk by Slidesgo (Autoguardado)Documento37 páginasEuropean Day of The Prevention of The Cardiovascular Risk by Slidesgo (Autoguardado)Myrian SantacruzAún no hay calificaciones
- Fisiología Endocrinologica IDocumento7 páginasFisiología Endocrinologica IMelissa sosa julcaAún no hay calificaciones
- Lab1 EndoDocumento20 páginasLab1 EndoJoshua MenesesAún no hay calificaciones
- Sistema EndocrinoDocumento9 páginasSistema EndocrinoMADDYAún no hay calificaciones
- Cap 13 Sistema EndócrinoDocumento10 páginasCap 13 Sistema Endócrinorodrigo4590Aún no hay calificaciones
- Sistema endocrino: glándulas y hormonasDocumento4 páginasSistema endocrino: glándulas y hormonasIng. Ludwing WG SanchezAún no hay calificaciones
- Sistema EndocrionoDocumento96 páginasSistema EndocrionoTalita IstefaneAún no hay calificaciones
- Compendio Examen Psicofisiología.Documento268 páginasCompendio Examen Psicofisiología.Maritza ArmijosAún no hay calificaciones
- Sistema Endocrino - HistoDocumento11 páginasSistema Endocrino - HistoLucas CordobaAún no hay calificaciones
- HipofisisDocumento9 páginasHipofisisAracely Torres ParraAún no hay calificaciones
- MarazDocumento39 páginasMarazWilber MarazAún no hay calificaciones
- Histo SeDocumento4 páginasHisto SeMin YoongiAún no hay calificaciones
- wuolah-free-TEMA-12-SISTEMA-ENDOCRINO-gulag-freeDocumento22 páginaswuolah-free-TEMA-12-SISTEMA-ENDOCRINO-gulag-freeUliAún no hay calificaciones
- Sistema EndocrinoDocumento7 páginasSistema EndocrinoPilarAún no hay calificaciones
- Glandulas EndocrinasDocumento21 páginasGlandulas EndocrinasHenry S. Carrillo G. DavilaAún no hay calificaciones
- Af1-Mod 6 - EndDocumento21 páginasAf1-Mod 6 - EndGustavo CamaraAún no hay calificaciones
- Mentefacto ConceptualDocumento7 páginasMentefacto ConceptualDaniel AriasAún no hay calificaciones
- Histo 2, Resumen Final, LabDocumento21 páginasHisto 2, Resumen Final, LabYORKIS SANTANA PEREZAún no hay calificaciones
- Sistema Endocrino GUIDEDocumento5 páginasSistema Endocrino GUIDEGabJimAún no hay calificaciones
- HipófisisDocumento2 páginasHipófisisZkϊppy 21Aún no hay calificaciones
- Histologia Organografica Endocrino 2023Documento89 páginasHistologia Organografica Endocrino 2023Bladimir CastroAún no hay calificaciones
- Sistema EndocrinoDocumento7 páginasSistema EndocrinoTomás JofreAún no hay calificaciones
- Teo 14. Sistema Endocrino 2017-1Documento71 páginasTeo 14. Sistema Endocrino 2017-1The goyos Aranda TrigosoAún no hay calificaciones
- Exposicion Unidad 2Documento32 páginasExposicion Unidad 2trabajosricardogaryAún no hay calificaciones
- Sistema EndocrinoDocumento5 páginasSistema EndocrinoJorgelis CuevasAún no hay calificaciones
- Sistema Endocrino22Documento142 páginasSistema Endocrino22Monse AlfaroAún no hay calificaciones
- Cap 21sistema EndocrinoHistoDocumento15 páginasCap 21sistema EndocrinoHistoPaolaAún no hay calificaciones
- Trabajo EndocrinoDocumento17 páginasTrabajo EndocrinoNathalia LopezAún no hay calificaciones
- Histo Ii. Semana 10. Teoria. Siste EndocrinoDocumento41 páginasHisto Ii. Semana 10. Teoria. Siste EndocrinoVictor Gavidia CaceresAún no hay calificaciones
- Sistema Endocrino (HISTOLOGIA) (Reydel Gonzalez)Documento5 páginasSistema Endocrino (HISTOLOGIA) (Reydel Gonzalez)Reydel Gonzalez RodriguezAún no hay calificaciones
- Sistemas endocrinos: Tejidos y hormonas claveDocumento11 páginasSistemas endocrinos: Tejidos y hormonas claveShei MendozaAún no hay calificaciones
- Sistema EndocrinoDocumento3 páginasSistema EndocrinoLuna GómezAún no hay calificaciones
- Sistema Endocrino IDocumento42 páginasSistema Endocrino IRuan Alves de AraujoAún no hay calificaciones
- Mis Notas de Histologia Sistema DigestivoDocumento9 páginasMis Notas de Histologia Sistema DigestivoTania VelázquezAún no hay calificaciones
- BIOENERGÉTICADocumento2 páginasBIOENERGÉTICATania VelázquezAún no hay calificaciones
- AGUADocumento2 páginasAGUATania VelázquezAún no hay calificaciones
- Aparato de GolgiDocumento2 páginasAparato de GolgiTania VelázquezAún no hay calificaciones
- 4Documento4 páginas4Tania VelázquezAún no hay calificaciones
- Sist Endocrino GuytonDocumento12 páginasSist Endocrino GuytonTania VelázquezAún no hay calificaciones
- Taller Hipotálamo HipófisisDocumento3 páginasTaller Hipotálamo HipófisisNoah Annelise OrtizAún no hay calificaciones
- Exposición Del DiencéfaloDocumento17 páginasExposición Del DiencéfaloRenzo ValverdeAún no hay calificaciones
- Asignacion Medicamentos EsteroidesDocumento4 páginasAsignacion Medicamentos Esteroidesjosefina diazAún no hay calificaciones
- PubertadDocumento6 páginasPubertadmisaelAún no hay calificaciones
- Triptico Aparato Reproductor Femenino y MasculinoDocumento3 páginasTriptico Aparato Reproductor Femenino y MasculinodannyAún no hay calificaciones
- CyD UP10 2022Documento52 páginasCyD UP10 2022Gabriela Delgado CastilloAún no hay calificaciones
- Apuntes HistologíaDocumento30 páginasApuntes HistologíaIngrid TomalaAún no hay calificaciones
- Órganos endocrinos y glándula tiroidesDocumento33 páginasÓrganos endocrinos y glándula tiroidesjohana paola merlano sotoAún no hay calificaciones
- Tarea 10.5 de AnatomiaDocumento4 páginasTarea 10.5 de AnatomiaRomero Marte hernandezAún no hay calificaciones
- Lesiones de La Piel 244444 Downloable 2060122Documento1 páginaLesiones de La Piel 244444 Downloable 2060122joanAún no hay calificaciones
- Pubertad Normal y Alteraciones Agosto 2016Documento49 páginasPubertad Normal y Alteraciones Agosto 2016Adela Marìa P LAún no hay calificaciones
- Mapa Conceptual 2Documento1 páginaMapa Conceptual 2Eduardo BritoAún no hay calificaciones
- Sistema Endocrino para Quinto de Primaria PDFDocumento5 páginasSistema Endocrino para Quinto de Primaria PDFLuzedy QuispeAún no hay calificaciones
- 2.1 Insulinoterapia 2019 PDFDocumento48 páginas2.1 Insulinoterapia 2019 PDFBenjamin Bastidas QuispecahuanaAún no hay calificaciones
- TiroidesDocumento27 páginasTiroidesMaría AlejandrinaAún no hay calificaciones
- Hipoglucemia y Administracion de GlucagonDocumento12 páginasHipoglucemia y Administracion de GlucagonManuel Tornero SoleraAún no hay calificaciones
- Cambios fisiológicos durante el embarazoDocumento12 páginasCambios fisiológicos durante el embarazowpaf234776Aún no hay calificaciones
- Hipotiroidismo e Hipertiroidismo.Documento42 páginasHipotiroidismo e Hipertiroidismo.Angelica100% (2)
- Ciclo Sexual Femenino 1221175409619246 9 PPT Share)Documento28 páginasCiclo Sexual Femenino 1221175409619246 9 PPT Share)DANIEL ALEJANDROAún no hay calificaciones
- Sistema Reproductor MasculinoDocumento35 páginasSistema Reproductor Masculinomonica rodriguez m100% (2)
- Fisiología de La LactanciaDocumento12 páginasFisiología de La LactanciaAnonymous GhJKML5CAún no hay calificaciones