También podría gustarte
- Crestas NeuralesDocumento2 páginasCrestas NeuralesOctavio ZmAún no hay calificaciones
- Crestas NeuralesDocumento2 páginasCrestas NeuralesOctavio ZmAún no hay calificaciones
- 3er Examen de PatologíaDocumento2 páginas3er Examen de PatologíaOctavio ZmAún no hay calificaciones
- Romero, Micología.Documento34 páginasRomero, Micología.Octavio ZmAún no hay calificaciones
- 4 - Reconocimiento Inmune Innato PDFDocumento5 páginas4 - Reconocimiento Inmune Innato PDFOctavio ZmAún no hay calificaciones
- IntroducciónDocumento5 páginasIntroducciónOctavio ZmAún no hay calificaciones
- AsfixiaDocumento72 páginasAsfixiaOctavio ZmAún no hay calificaciones
- Romero Generalidades PDFDocumento10 páginasRomero Generalidades PDFOctavio ZmAún no hay calificaciones
- AntroDocumento2 páginasAntroOctavio ZmAún no hay calificaciones
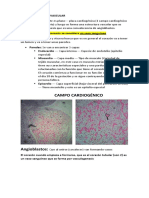
- Circulación CoronariaDocumento13 páginasCirculación CoronariaOctavio ZmAún no hay calificaciones
- Parasitología: Generalidades y protozoos comensalesDocumento65 páginasParasitología: Generalidades y protozoos comensalesadriansotegarAún no hay calificaciones
- BacteriologiaDocumento78 páginasBacteriologiaFany Espejel FigueroaAún no hay calificaciones
- Inmunodeficiencias Primarias PDFDocumento6 páginasInmunodeficiencias Primarias PDFOctavio ZmAún no hay calificaciones
- VACUNACIÓNDocumento4 páginasVACUNACIÓNNoemi MccartneyAún no hay calificaciones
- 3 - Barreras Físicas, Químicas y Microbiológicas PDFDocumento2 páginas3 - Barreras Físicas, Químicas y Microbiológicas PDFOctavio ZmAún no hay calificaciones
- Complejo Mayor de Histocompatibilidad PDFDocumento5 páginasComplejo Mayor de Histocompatibilidad PDFOctavio Zm0% (1)
- Inflamación y FagocitosisDocumento4 páginasInflamación y FagocitosisOctavio ZmAún no hay calificaciones
- TransplanteDocumento4 páginasTransplanteOctavio ZmAún no hay calificaciones
- Subpoblaciones de T CooperadoresDocumento3 páginasSubpoblaciones de T CooperadoresOctavio ZmAún no hay calificaciones
- Inmunodeficiencias: causas, tipos y efectosDocumento4 páginasInmunodeficiencias: causas, tipos y efectosOctavio ZmAún no hay calificaciones
- Inmunidad frente a tumoresDocumento4 páginasInmunidad frente a tumoresOctavio ZmAún no hay calificaciones
- Tolerancia y Autoinmunidad PDFDocumento5 páginasTolerancia y Autoinmunidad PDFOctavio ZmAún no hay calificaciones
- Inmunidad A PatógenosDocumento6 páginasInmunidad A PatógenosOctavio ZmAún no hay calificaciones
- InmunologíaDocumento2 páginasInmunologíaOctavio ZmAún no hay calificaciones
- InmunopatologíaDocumento7 páginasInmunopatologíaalfredoAún no hay calificaciones
- CITOTOXICIDAD NK Y CTLDocumento2 páginasCITOTOXICIDAD NK Y CTLOctavio ZmAún no hay calificaciones
- Subpoblaciones de T Cooperadores PDFDocumento3 páginasSubpoblaciones de T Cooperadores PDFOctavio ZmAún no hay calificaciones
- Citocinas PDFDocumento5 páginasCitocinas PDFCris Dev100% (1)
- BCR y Activación Del BDocumento2 páginasBCR y Activación Del Bcharlie.9411Aún no hay calificaciones
- Abreviaturas de InmunologíaDocumento2 páginasAbreviaturas de InmunologíaMichelle Dl Angeles Suarez67% (6)
- Sistemas de defensa del organismoDocumento22 páginasSistemas de defensa del organismoKarla BastidaAún no hay calificaciones
- Celulas NKDocumento8 páginasCelulas NKHelen ZambranoAún no hay calificaciones
- Final IiDocumento6 páginasFinal IiLetícia Raquel PintoAún no hay calificaciones
- Licenciatura en BiotecnologiaDocumento29 páginasLicenciatura en BiotecnologiaMartina ColmegnaAún no hay calificaciones
- Virologia IntroduccionDocumento84 páginasVirologia IntroduccionAnnie MomAún no hay calificaciones
- Infografia Linfocito TDocumento1 páginaInfografia Linfocito TDaniela FloresAún no hay calificaciones
- Inmunidad a mucosas: Tejido linfoide asociado y respuestas innatasDocumento74 páginasInmunidad a mucosas: Tejido linfoide asociado y respuestas innatasVanessa YapoAún no hay calificaciones
- Línea eritroide: diferenciación y maduración de los precursores eritocitariosDocumento20 páginasLínea eritroide: diferenciación y maduración de los precursores eritocitariosRocioAún no hay calificaciones
- Resumen Inmunologia Teorico 2020Documento158 páginasResumen Inmunologia Teorico 2020Romy PechAún no hay calificaciones
- Resistencia Del Organismo A La Infección II: Inmunidad y AlergiaDocumento28 páginasResistencia Del Organismo A La Infección II: Inmunidad y Alergiafelix GarcíaAún no hay calificaciones
- Manual de Vacunas de Latinoamerica Ed 2021Documento750 páginasManual de Vacunas de Latinoamerica Ed 2021jaradiamax100% (1)
- INMUNOFARMACOLOGIA DiaposDocumento31 páginasINMUNOFARMACOLOGIA DiaposAnonymous 8MDxKs0% (1)
- Dermatomiositis, polimiositis y esclerosis sistémica progresivaDocumento36 páginasDermatomiositis, polimiositis y esclerosis sistémica progresivaDeltaScarlettFigueraAún no hay calificaciones
- Complejo Principal de HistocompatibilidadDocumento8 páginasComplejo Principal de HistocompatibilidadluisAún no hay calificaciones
- Células Del Sistema InmunitarioDocumento10 páginasCélulas Del Sistema InmunitarioReDaZuKa100% (3)
- Caracteristicas Principales de EritrocitosDocumento7 páginasCaracteristicas Principales de EritrocitosJeaneth Peñaranda AranibarAún no hay calificaciones
- InmunopatologiaDocumento3 páginasInmunopatologiaMaria Jose ViloriaAún no hay calificaciones
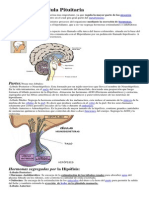
- Glándula Pituitaria y sus FuncionesDocumento13 páginasGlándula Pituitaria y sus FuncionesPia Dominguez AlzagaAún no hay calificaciones
- Respuesta Inmune DengueDocumento5 páginasRespuesta Inmune DengueAnonymous ApyNcvV2VlAún no hay calificaciones
- Inmunologia de La EclampsiaDocumento8 páginasInmunologia de La EclampsiaMarco Carbonel100% (1)
- 2019 Revision Literatura VirusDocumento8 páginas2019 Revision Literatura VirusHECTOR ELBER CAMACHO GUTIERREZAún no hay calificaciones
- Tema 12 Sistema Inmune BIOLOGÍADocumento15 páginasTema 12 Sistema Inmune BIOLOGÍApatricia mata moyaAún no hay calificaciones
- Ensayo sobre los tipos de ensayo y sus característicasDocumento5 páginasEnsayo sobre los tipos de ensayo y sus característicasCristina EspinosaAún no hay calificaciones
- Generalidades de InmunologiaDocumento47 páginasGeneralidades de InmunologiaAnonymous CoUBbG1mLAún no hay calificaciones
- Derrame Pleural Tuberculoso.Documento9 páginasDerrame Pleural Tuberculoso.Yony Morales LeonAún no hay calificaciones
- 1 Corte Histo DefinitivoDocumento62 páginas1 Corte Histo DefinitivoCESAR AUGUSTO ARIAS MOLANOAún no hay calificaciones
- Test de AdaDocumento10 páginasTest de AdaPaúlEduardoCervantesParvinaAún no hay calificaciones
- Resumen 4 CapsDocumento37 páginasResumen 4 CapsdaniaAún no hay calificaciones
- Monografia Del Reconocimiento de Órganos Linfoides en AvesDocumento14 páginasMonografia Del Reconocimiento de Órganos Linfoides en AvesCristobal Severo Huamani Carrion100% (4)