También podría gustarte
- Integración Del Metabolismo IIIDocumento5 páginasIntegración Del Metabolismo IIIJavier MendozaAún no hay calificaciones
- Bioquimica de Tejidos (Blanco)Documento35 páginasBioquimica de Tejidos (Blanco)Francisco ReyesAún no hay calificaciones
- X9 Metabolismo de Carbohidratos PDFDocumento15 páginasX9 Metabolismo de Carbohidratos PDFCote VegaAún no hay calificaciones
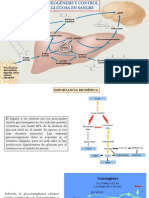
- Gluconeogénesis, Glucogenogénesis y GlucogenólisisDocumento80 páginasGluconeogénesis, Glucogenogénesis y GlucogenólisisValdovinos KennethAún no hay calificaciones
- Bioquimica Barcelo Actividad Obligatoria - Ud7 - Tema 1Documento3 páginasBioquimica Barcelo Actividad Obligatoria - Ud7 - Tema 1carlos villamonte100% (7)
- Metabolismo de Los Hidratos de CarbonoDocumento38 páginasMetabolismo de Los Hidratos de CarbonoJosé Fernando López JiménezAún no hay calificaciones
- GluconeogenesisDocumento9 páginasGluconeogenesisAyeleen Alvarado gallegosAún no hay calificaciones
- El metabolismo de los carbohidratos: las principales vías de digestión, transporte y degradaciónDocumento35 páginasEl metabolismo de los carbohidratos: las principales vías de digestión, transporte y degradaciónJoelTevesMendoza100% (2)
- Función HepáticaDocumento5 páginasFunción HepáticaDemci Mendoza0% (1)
- Tem 6 BioqDocumento10 páginasTem 6 BioqAlexandra María Marín BernalAún no hay calificaciones
- Gluconeogénesis: Síntesis de glucosa a partir de otros sustratosDocumento18 páginasGluconeogénesis: Síntesis de glucosa a partir de otros sustratosCarlota GuardiaAún no hay calificaciones
- Generalidades Del MetabolismoDocumento19 páginasGeneralidades Del MetabolismoCarlos ChiaAún no hay calificaciones
- Metabolismo Glucólisis (ALUMNOS 3)Documento22 páginasMetabolismo Glucólisis (ALUMNOS 3)danielaestefaniasalinasrojas8Aún no hay calificaciones
- 4-Integracion Metabolica - Parte B AyunoDocumento7 páginas4-Integracion Metabolica - Parte B AyunoMailén GajewskiAún no hay calificaciones
- UABP 3. Metabolismo en Algunos TejidosDocumento8 páginasUABP 3. Metabolismo en Algunos TejidosMartín PanicoAún no hay calificaciones
- Transportadores de GlucosaDocumento5 páginasTransportadores de GlucosaHansel DueñasAún no hay calificaciones
- Trabajo Practico Nº6Documento6 páginasTrabajo Practico Nº6Jose Ignacio BarrientosAún no hay calificaciones
- Hormona GlucagonDocumento6 páginasHormona GlucagonNicolásAún no hay calificaciones
- Chopita - Regulacion, Integracion Metabolica y Enfermedades TEXTUAL AL TEORICODocumento5 páginasChopita - Regulacion, Integracion Metabolica y Enfermedades TEXTUAL AL TEORICOCarolina PeralesAún no hay calificaciones
- Macronutrientes SSDocumento69 páginasMacronutrientes SSJULIETAAún no hay calificaciones
- RUTA METABÓLICA ExposicionDocumento27 páginasRUTA METABÓLICA Exposicionduber chonlon guevaraAún no hay calificaciones
- BIOQUIMICADocumento15 páginasBIOQUIMICAADRIANA PAOLA CUESTA GARCÉSAún no hay calificaciones
- Integración Metabólica - ResumenDocumento10 páginasIntegración Metabólica - ResumenJulianne RestrepoAún no hay calificaciones
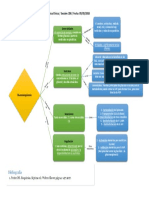
- Mapa Conceptual Acerca Del Metabolismo de GluconeogénesisDocumento1 páginaMapa Conceptual Acerca Del Metabolismo de GluconeogénesisMauricio Caña Avalos89% (9)
- Control 3 BioquímicaDocumento7 páginasControl 3 BioquímicaJaviera ArandaAún no hay calificaciones
- GluconeogenesisDocumento2 páginasGluconeogenesisAnnyVal RMAún no hay calificaciones
- GLUCONEOGENESISSDocumento23 páginasGLUCONEOGENESISSJaviAún no hay calificaciones
- Metabolismo de Los GlúcidosDocumento10 páginasMetabolismo de Los GlúcidosAguus CabreraAún no hay calificaciones
- Wuolah Free Bloque 6Documento10 páginasWuolah Free Bloque 6jaja jajaAún no hay calificaciones
- GluconeogenesisDocumento45 páginasGluconeogenesisKBAún no hay calificaciones
- Gluconeogénesis: síntesis de glucosa a partir de precursores no glucídicosDocumento9 páginasGluconeogénesis: síntesis de glucosa a partir de precursores no glucídicosBerta González GómezAún no hay calificaciones
- Integración Metabólica en El Estado PostabsortivoDocumento2 páginasIntegración Metabólica en El Estado PostabsortivoangelikAún no hay calificaciones
- Funcion HepaticaDocumento58 páginasFuncion HepaticaYamili Condori100% (1)
- Bioka Sem Expo ErikaDocumento8 páginasBioka Sem Expo ErikaErika Ruiz TorresAún no hay calificaciones
- GluconeogénesisDocumento16 páginasGluconeogénesisNATALIA DANIELA VILLADIEGO BERROCALAún no hay calificaciones
- CARBOHIDRATOS UNIFICADOoDocumento11 páginasCARBOHIDRATOS UNIFICADOoJesus Smaikel Teran MuenalaAún no hay calificaciones
- Fisiología de SistemasDocumento20 páginasFisiología de SistemasLaura MenaAún no hay calificaciones
- Regulacion Hormonal Del Metabolismo EnergeticoDocumento8 páginasRegulacion Hormonal Del Metabolismo EnergeticoDaniAún no hay calificaciones
- Metabolismo Energético y Homeostasis de La GlucosaDocumento6 páginasMetabolismo Energético y Homeostasis de La GlucosaLuana LAún no hay calificaciones
- Gluconeogenesis y Control de La Glucosa en SangreDocumento37 páginasGluconeogenesis y Control de La Glucosa en SangreStefano GarciaAún no hay calificaciones
- Bioquímica Mckee. Metabolismo de CarbohidratosDocumento38 páginasBioquímica Mckee. Metabolismo de CarbohidratosNancy CarolinaAún no hay calificaciones
- Entrada de GlucosaDocumento4 páginasEntrada de GlucosaLourdes PerdomoAún no hay calificaciones
- GLUCONEOGÉNESISDocumento22 páginasGLUCONEOGÉNESISyuri yuriAún no hay calificaciones
- GluconeogénesisDocumento40 páginasGluconeogénesisBritez Paredes Alejandra SilvanaAún no hay calificaciones
- Metabolismo proteico: equilibrio entre síntesis y degradaciónDocumento3 páginasMetabolismo proteico: equilibrio entre síntesis y degradaciónRosario BarrientosAún no hay calificaciones
- TEMA 02 - Metabolismo de Tejidos y ÓrganosDocumento16 páginasTEMA 02 - Metabolismo de Tejidos y ÓrganosDavid García MartínezAún no hay calificaciones
- Integracion MetabolicaDocumento12 páginasIntegracion MetabolicaFlor SternariAún no hay calificaciones
- Respuesta Metabólica Al Estrés Quirúrgico (Material para Estudiantes) .Documento18 páginasRespuesta Metabólica Al Estrés Quirúrgico (Material para Estudiantes) .MARIA GABRIELA HERNANDEZ GONTOLAún no hay calificaciones
- MATEMÁTICADocumento3 páginasMATEMÁTICADomenica Maribel Zambrano CedeñoAún no hay calificaciones
- Lectura - Metabolismo de Los CarbohidratosDocumento24 páginasLectura - Metabolismo de Los CarbohidratosFernando Naim Inga MorenoAún no hay calificaciones
- Glicólisis EVEADocumento76 páginasGlicólisis EVEASusanaAún no hay calificaciones
- Clase 2 FseDocumento13 páginasClase 2 FseAIXIA ZARRICUETA PERALTAAún no hay calificaciones
- Metabolismo Del Higado 1Documento26 páginasMetabolismo Del Higado 1gustavo gaitanAún no hay calificaciones
- Tema 16Documento7 páginasTema 16IvanaAún no hay calificaciones
- Apuntes Tema 5 - Alteraciones Del Metabolismo de Los GlúcidosDocumento16 páginasApuntes Tema 5 - Alteraciones Del Metabolismo de Los GlúcidosChristian García TorresAún no hay calificaciones
- La gluconeogénesis, clave para la homeostasis glucémicaDocumento5 páginasLa gluconeogénesis, clave para la homeostasis glucémicaEl club de la LecturaAún no hay calificaciones
- Cap 68 GUYTONDocumento4 páginasCap 68 GUYTONRosario Barrientos100% (1)
- Unidada 5. Metabolismo HdeC - GlicolisisDocumento29 páginasUnidada 5. Metabolismo HdeC - GlicolisisMaximimilianoAún no hay calificaciones
- Mi Resumen MIR EndocrinologíaDocumento56 páginasMi Resumen MIR Endocrinologíaluis filosAún no hay calificaciones
- Resumen Protocolo de KiotoDocumento1 páginaResumen Protocolo de KiotoDenisseRamosHernndezAún no hay calificaciones
- Textos PDFDocumento4 páginasTextos PDFJuan Pablo Martínez GómezAún no hay calificaciones
- Lista de Verificación de Desarrollo de Motricidad FinaDocumento6 páginasLista de Verificación de Desarrollo de Motricidad FinaKathe Londoño CAún no hay calificaciones
- Juegos para pasar el verano en familiaDocumento61 páginasJuegos para pasar el verano en familiaUniformes AdrianaAún no hay calificaciones
- JorgeMendietaMuñoz Tarea 1Documento16 páginasJorgeMendietaMuñoz Tarea 1yuli yimena rueda tellezAún no hay calificaciones
- Analisis de Sedimentos en La CaptacionDocumento30 páginasAnalisis de Sedimentos en La Captacionyude magali lopez arvaloAún no hay calificaciones
- Análisis de RegresiónDocumento23 páginasAnálisis de RegresiónANGHELO STEFFANO PAREDES GUTIERREZAún no hay calificaciones
- La Raíz BotanicaDocumento55 páginasLa Raíz Botanicaselvin100% (2)
- Astra GDocumento4 páginasAstra GDaniel RedondoAún no hay calificaciones
- Control de Generacion de Gases Toxicos en Voladuras en Minas A Cielo Abierto PDFDocumento15 páginasControl de Generacion de Gases Toxicos en Voladuras en Minas A Cielo Abierto PDFJose RojasAún no hay calificaciones
- Calendario OMSDocumento12 páginasCalendario OMSJulia Torres AcostaAún no hay calificaciones
- Evolución Del Derecho Positivo Peruano Código Civil 1852 y Tratado de Lima 1877Documento5 páginasEvolución Del Derecho Positivo Peruano Código Civil 1852 y Tratado de Lima 1877raul rodolfo oregon palominoAún no hay calificaciones
- Ficha Tecnica Excavadora ZX120Documento4 páginasFicha Tecnica Excavadora ZX120ferchoAún no hay calificaciones
- Elaboración Del Plan de Mantenimiento para La Linea de Producción Del Ladrillo en La Empresa Ladrillera Arcillas LtdaDocumento166 páginasElaboración Del Plan de Mantenimiento para La Linea de Producción Del Ladrillo en La Empresa Ladrillera Arcillas Ltdahumberto quamiAún no hay calificaciones
- Examen Final de Trabajo en Altura - Revisión Del IntentoDocumento6 páginasExamen Final de Trabajo en Altura - Revisión Del Intentocachupin12100% (2)
- ACT 1 - Castro GiulianaDocumento5 páginasACT 1 - Castro GiulianaGiuliana CastroAún no hay calificaciones
- Art 3bbbDocumento18 páginasArt 3bbbwackkofranAún no hay calificaciones
- Viaje A La Luna (Información)Documento15 páginasViaje A La Luna (Información)3M1N3MAún no hay calificaciones
- Lecciones Del Huevo CompletoDocumento176 páginasLecciones Del Huevo Completodaviesgay50% (2)
- TEST COMPRENSIVO FiloDocumento2 páginasTEST COMPRENSIVO FiloLuis Fernando Duran PereaAún no hay calificaciones
- NOCTURNAL Total La Distancia DetallesDocumento5 páginasNOCTURNAL Total La Distancia DetallesDiarios camAún no hay calificaciones
- Como Se Clasifican Las Áreas Eléctricas Peligrosas Según La NEC PDFDocumento7 páginasComo Se Clasifican Las Áreas Eléctricas Peligrosas Según La NEC PDFguillermo uribeAún no hay calificaciones
- Etica Profesional PDFDocumento9 páginasEtica Profesional PDFAngelKBattlerAún no hay calificaciones
- Preoperacional de Equipo de Soldar..Documento1 páginaPreoperacional de Equipo de Soldar..Juan PayaresAún no hay calificaciones
- Septiembre AlumnosDocumento4 páginasSeptiembre AlumnosManuel Muriel FloresAún no hay calificaciones
- Pruebas Católica ContabilidadDocumento61 páginasPruebas Católica ContabilidadJose Fernando Acuña QuirogaAún no hay calificaciones
- Creación de Cubos de Datos Mediante Archivos de Excel y de TDocumento8 páginasCreación de Cubos de Datos Mediante Archivos de Excel y de THarold OrtegaAún no hay calificaciones
- C11-EBRS-22 EBR Secundaria Ciencia y Tecnología PDFDocumento67 páginasC11-EBRS-22 EBR Secundaria Ciencia y Tecnología PDFHector Moises Baes VillafuerteAún no hay calificaciones
- Informe DETERMINACION DE LA LECHE.Documento12 páginasInforme DETERMINACION DE LA LECHE.jose HERNANDEZAún no hay calificaciones
- 09 PRACTICA PRUEBA HIPOTESIS PROPORCIONES EstudDocumento5 páginas09 PRACTICA PRUEBA HIPOTESIS PROPORCIONES EstudpaolaAún no hay calificaciones