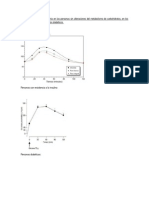
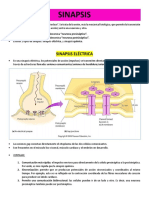
También podría gustarte
- Cuestionario 3Documento2 páginasCuestionario 3luciasc93100% (7)
- Informe 6 Bioquímica SeminarioDocumento12 páginasInforme 6 Bioquímica SeminarioEmery Sofía C. D. BustamanteAún no hay calificaciones
- Recepción CelularDocumento11 páginasRecepción CelularStephano P Carranza75% (4)
- Factores que influyen en lípidos séricosDocumento3 páginasFactores que influyen en lípidos séricosLucia Saucedo TalledoAún no hay calificaciones
- Enzimas necesarias para que la levadura aproveche el almidónDocumento2 páginasEnzimas necesarias para que la levadura aproveche el almidónXiomara Janne Linares SaldañaAún no hay calificaciones
- Mecanismos AutocontrolDocumento4 páginasMecanismos AutocontrolLuis Jara Vidalon OrrilloAún no hay calificaciones
- Cuestionario 8Documento3 páginasCuestionario 8Katherine Ramos Calderón50% (2)
- Informe P3, Kmalpartidad-06t10-1Documento7 páginasInforme P3, Kmalpartidad-06t10-1Johnattan Javier Espinoza Castañeda0% (1)
- Informe Fisica 9Documento10 páginasInforme Fisica 9Angelina100% (1)
- Exposicion S9 - Fisiopato Seminario - Grupo Es029 - 1Documento34 páginasExposicion S9 - Fisiopato Seminario - Grupo Es029 - 1Brian linares RodriguezAún no hay calificaciones
- Informe 2Documento3 páginasInforme 2Milene QAún no hay calificaciones
- Informe 10 Bioquímica LabDocumento3 páginasInforme 10 Bioquímica LabRayza Pomatay VasquezAún no hay calificaciones
- Tarea 6 - Grupo 21B - Bioquímica SeminarioDocumento12 páginasTarea 6 - Grupo 21B - Bioquímica SeminarioGuianela Polo100% (1)
- Biologia SEMANA 13Documento3 páginasBiologia SEMANA 13john salchichon100% (5)
- Estructuras y Funciones Normales Del Gen Ada y Su ProteínaDocumento1 páginaEstructuras y Funciones Normales Del Gen Ada y Su ProteínaMelanyAún no hay calificaciones
- Pruebas No Paramétricas Aplicadas A La Medicina: Mann Whitney, WilcoxonDocumento5 páginasPruebas No Paramétricas Aplicadas A La Medicina: Mann Whitney, WilcoxonChristianPLAYAún no hay calificaciones
- Anatomia Informe 6Documento13 páginasAnatomia Informe 6luiguiman123456Aún no hay calificaciones
- Tarea 13 - Grupo 25aDocumento10 páginasTarea 13 - Grupo 25aChristianPLAYAún no hay calificaciones
- Examenes Auxiliares - DBM2Documento6 páginasExamenes Auxiliares - DBM2Reynaldo Vasquez GarciaAún no hay calificaciones
- Detección actividad enzimas catalasa amilasaDocumento12 páginasDetección actividad enzimas catalasa amilasaSergio Leonel Moran Stechmann0% (1)
- Pregunta 4 y 5Documento3 páginasPregunta 4 y 5claudia barrantes castilloAún no hay calificaciones
- Sseminario 9 Recepción CelularDocumento1 páginaSseminario 9 Recepción CelularAnonymous hkjp4qMVRsAún no hay calificaciones
- Informe 9 Micro PracDocumento6 páginasInforme 9 Micro PracAlexa OréAún no hay calificaciones
- Exposición S3 - Informática II - Racarleyl-3Documento38 páginasExposición S3 - Informática II - Racarleyl-3Luciana Rivasplata CastroAún no hay calificaciones
- Mapa Conceptual Seminario 12Documento1 páginaMapa Conceptual Seminario 12Fabrizzio Morales Zaldivar100% (1)
- Informe S6 - Fisiología PrácticaDocumento10 páginasInforme S6 - Fisiología PrácticaCoraine Chávez CórdovaAún no hay calificaciones
- Fisiolab GlicemiaDocumento3 páginasFisiolab GlicemiaEstela ChumbirayAún no hay calificaciones
- Práctica+ ElectriDocumento21 páginasPráctica+ ElectriJose Jhon Suni Aslla50% (4)
- Un MultitésterDocumento2 páginasUn MultitésterDiana Iparraguirre100% (1)
- METABOLISMO DE CARBOHIDRATOS EN LAS FASES POSTPRANDIAL, POSTABSORTIVA Y AYUNODocumento12 páginasMETABOLISMO DE CARBOHIDRATOS EN LAS FASES POSTPRANDIAL, POSTABSORTIVA Y AYUNOEmery Sofía C. D. Bustamante0% (1)
- Mediciones e Indicadores EpidemiológicasDocumento14 páginasMediciones e Indicadores EpidemiológicasvibescashAún no hay calificaciones
- Búsqueda Con Descriptores y Clasificación de Resultados en Zotero (Avelasquezj-16)Documento35 páginasBúsqueda Con Descriptores y Clasificación de Resultados en Zotero (Avelasquezj-16)salma pejerreyAún no hay calificaciones
- Deposito Extracelular y Alteración Crecimiento y DesarrolloDocumento6 páginasDeposito Extracelular y Alteración Crecimiento y DesarrolloStephano Ramirez0% (1)
- Informe S1 - Grupo 11 - Integración de Los Fenómenos Eléctricos, Mecánicos, Hemodinámicos y Sonoros Del Corazón - FISIOLOGÍA SEMINARIODocumento22 páginasInforme S1 - Grupo 11 - Integración de Los Fenómenos Eléctricos, Mecánicos, Hemodinámicos y Sonoros Del Corazón - FISIOLOGÍA SEMINARIOvibescash100% (1)
- Informe 3 - Fisiopatologia SeminarioDocumento19 páginasInforme 3 - Fisiopatologia SeminarioMilagros Bustamante100% (1)
- Biologia SEMANA 11Documento3 páginasBiologia SEMANA 11john salchichon90% (10)
- Experimento 2Documento3 páginasExperimento 2CarlosSonoAún no hay calificaciones
- Fisiología Práctica Informe 8Documento4 páginasFisiología Práctica Informe 8Rayza Pomatay Vasquez100% (1)
- Informática - Semana 06Documento4 páginasInformática - Semana 06Erick TelloAún no hay calificaciones
- Informe 10 - Grupo 10-Fisiopatologia SeminarioDocumento7 páginasInforme 10 - Grupo 10-Fisiopatologia SeminarioViviana LopezAún no hay calificaciones
- Tarea 11 Fisica BioelectrecidadDocumento6 páginasTarea 11 Fisica BioelectrecidadCeleste VásquezAún no hay calificaciones
- Moléculas de Relación CelularDocumento15 páginasMoléculas de Relación CelularAlejandro SuaresAún no hay calificaciones
- Cuestionario-14 VIRUSDocumento6 páginasCuestionario-14 VIRUSJheanPaulAún no hay calificaciones
- Bio Sem InformeDocumento15 páginasBio Sem InformeXiomara Janne Linares SaldañaAún no hay calificaciones
- Hidrodinámica Médica IDocumento2 páginasHidrodinámica Médica IA.V.M.100% (6)
- Observacion de Embriones - Cavidad Abdominal. Aparato Renal LAB EMBRIODocumento11 páginasObservacion de Embriones - Cavidad Abdominal. Aparato Renal LAB EMBRIORenzo Enrique Verastegui BezzoloAún no hay calificaciones
- Exposición 04 - Farmacología Seminario - Es025Documento24 páginasExposición 04 - Farmacología Seminario - Es025Guianela PoloAún no hay calificaciones
- Seminario Fisiopatologia ComaDocumento36 páginasSeminario Fisiopatologia Comasergio leonel moran stechmann100% (1)
- ElectrocardiografiaDocumento5 páginasElectrocardiografiaSamer BastAún no hay calificaciones
- Famaco Lab Exposición 4Documento15 páginasFamaco Lab Exposición 4Edena Pérez Del ÁguilaAún no hay calificaciones
- Base fisiológica cambios observados experimentosDocumento1 páginaBase fisiológica cambios observados experimentosEricksen SalazarAún no hay calificaciones
- Informe 12Documento8 páginasInforme 12Marisabel GuerraAún no hay calificaciones
- Expo Sapu Sem 4Documento8 páginasExpo Sapu Sem 4sandraAún no hay calificaciones
- Filamentos celularesDocumento4 páginasFilamentos celularesKenner Poult Santisteban Muñoz100% (1)
- Dismenorrea AINEsDocumento21 páginasDismenorrea AINEsvibescashAún no hay calificaciones
- Fisiopatologìa Seminario Mapa 12Documento3 páginasFisiopatologìa Seminario Mapa 12Vanessa AvilaAún no hay calificaciones
- Acción de La Sustancia Transmisora en La Neurona PostsinápticaDocumento2 páginasAcción de La Sustancia Transmisora en La Neurona PostsinápticaJohnny Mauricio0% (1)
- SINAPSISDocumento56 páginasSINAPSISDomenica MeraAún no hay calificaciones
- Canales iónicos y farmacologíaDocumento27 páginasCanales iónicos y farmacologíalizbeth calderon acostaAún no hay calificaciones
- Sistema nervioso central: Transmisores y receptoresDocumento16 páginasSistema nervioso central: Transmisores y receptoresClaudia MaldonadoAún no hay calificaciones
- Generalidades 1 RX SMT 2020Documento98 páginasGeneralidades 1 RX SMT 2020Yakelin Villanueva100% (1)
- Imipenem-Cilastatina y MeropenemDocumento1 páginaImipenem-Cilastatina y MeropenemYakelin VillanuevaAún no hay calificaciones
- LupusDocumento11 páginasLupusYakelin VillanuevaAún no hay calificaciones
- Fisiopato 2019-2 SeminariosDocumento15 páginasFisiopato 2019-2 SeminariosCamila Rodriguez AgurtoAún no hay calificaciones
- AINES Estructura-ActividadDocumento21 páginasAINES Estructura-ActividadMaria Quiñones HuayaneyAún no hay calificaciones
- AteromaDocumento7 páginasAteromaYakelin VillanuevaAún no hay calificaciones
- Práctica 7 PulmonarDocumento2 páginasPráctica 7 PulmonarJesús MoraAún no hay calificaciones
- Trast. HemodinamicosDocumento56 páginasTrast. HemodinamicosYakelin VillanuevaAún no hay calificaciones
- Trastornos de CoagulaciónDocumento10 páginasTrastornos de CoagulaciónYakelin VillanuevaAún no hay calificaciones
- Beta Agonistas en El AsmaDocumento8 páginasBeta Agonistas en El AsmaYakelin VillanuevaAún no hay calificaciones
- Trastornos de CoagulaciónDocumento10 páginasTrastornos de CoagulaciónYakelin VillanuevaAún no hay calificaciones
- Fisiopatología de la diabetes y mecanismos de muerte de células βDocumento9 páginasFisiopatología de la diabetes y mecanismos de muerte de células βJimmy StephensAún no hay calificaciones
- ComaDocumento3 páginasComaYakelin VillanuevaAún no hay calificaciones
- Estados de ConcienciaDocumento9 páginasEstados de ConcienciaValeria CerdaAún no hay calificaciones
- Teorias PDFDocumento2 páginasTeorias PDFYakelin VillanuevaAún no hay calificaciones
- Articulo de HTA Fisiopatologia PDFDocumento18 páginasArticulo de HTA Fisiopatologia PDFMauricio Mormontoy ÑiqueAún no hay calificaciones
- AINES ScieloDocumento9 páginasAINES ScieloJuan Rodrigo Lisboa San MartínAún no hay calificaciones
- Coma 1Documento14 páginasComa 1Yakelin VillanuevaAún no hay calificaciones
- Guia Practica Patología I 2019-IiDocumento73 páginasGuia Practica Patología I 2019-IiYakelin VillanuevaAún no hay calificaciones
- Estados de ConcienciaDocumento9 páginasEstados de ConcienciaValeria CerdaAún no hay calificaciones
- Farmaco NeuroDocumento15 páginasFarmaco NeuroYakelin VillanuevaAún no hay calificaciones
- Sem 4 Lectura 1 Farmaco en Ansiedad - En.esDocumento14 páginasSem 4 Lectura 1 Farmaco en Ansiedad - En.esYakelin VillanuevaAún no hay calificaciones
- AINES ScieloDocumento9 páginasAINES ScieloJuan Rodrigo Lisboa San MartínAún no hay calificaciones
- Farmaco Sem 1Documento5 páginasFarmaco Sem 1Yakelin VillanuevaAún no hay calificaciones
- Enfermedad Vascular CerebralDocumento11 páginasEnfermedad Vascular Cerebralfrey7Aún no hay calificaciones
- Fisiología gastrointestinal y defecaciónDocumento106 páginasFisiología gastrointestinal y defecaciónYakelin VillanuevaAún no hay calificaciones
- Enzimas de Fase 2Documento1 páginaEnzimas de Fase 2Yakelin VillanuevaAún no hay calificaciones
- Ejer Cici OsDocumento9 páginasEjer Cici OsYakelin VillanuevaAún no hay calificaciones
- Caso Clínico N°1-Pregunta 2Documento5 páginasCaso Clínico N°1-Pregunta 2Yakelin VillanuevaAún no hay calificaciones
- Apuntes Ácido BaseDocumento4 páginasApuntes Ácido BaseYakelin VillanuevaAún no hay calificaciones
- Órgano Tendinoso de GolgiDocumento1 páginaÓrgano Tendinoso de Golgi05-OD-HU-DIONICIO SIERRA BERRIOAún no hay calificaciones
- Lab. Fisio. Prac. 5 FinalDocumento9 páginasLab. Fisio. Prac. 5 FinalGera HernandezAún no hay calificaciones
- NEUROFISIOLOGIADocumento17 páginasNEUROFISIOLOGIAEderOcampoPerez100% (2)
- Los Principales NeurotransmisoresDocumento9 páginasLos Principales NeurotransmisoresYadi QuiñonesAún no hay calificaciones
- TP 1 2020 Union NeuromuscularDocumento6 páginasTP 1 2020 Union NeuromuscularAgustina EcheverriaAún no hay calificaciones
- ABSTRACTO Neuroplasticidad Aspectos BioquímicosDocumento14 páginasABSTRACTO Neuroplasticidad Aspectos BioquímicosJose PichardoAún no hay calificaciones
- Reflejos monosinapticos y polisinapticos: estudio de respuestas muscularesDocumento7 páginasReflejos monosinapticos y polisinapticos: estudio de respuestas muscularesCristian Pertuz0% (1)
- Guia Sistema Nervios-SinapsisDocumento7 páginasGuia Sistema Nervios-SinapsisRec Gruas beltreAún no hay calificaciones
- Neurotransmisores y sinapsis: características y procesos claveDocumento7 páginasNeurotransmisores y sinapsis: características y procesos claveAustriaOoAún no hay calificaciones
- SinapsisDocumento4 páginasSinapsisCesar Duque50% (2)
- Neurofisiologia ReflejosDocumento23 páginasNeurofisiologia ReflejosAurora ChumpitazAún no hay calificaciones
- Comunicación Neuronal PDFDocumento1 páginaComunicación Neuronal PDFDavidAún no hay calificaciones
- Reflejos Y Niveles de Integración NerviosaDocumento28 páginasReflejos Y Niveles de Integración NerviosaSofia MartinezAún no hay calificaciones
- Taller Histofisiolofia Del Sistema Nervioso y NeurotransmisoresDocumento4 páginasTaller Histofisiolofia Del Sistema Nervioso y NeurotransmisoresAngie Nathalia Cutiva MedinaAún no hay calificaciones
- El OlvidoDocumento7 páginasEl OlvidoIrene Alvear AzcárateAún no hay calificaciones
- Ejercicios Neurotróficos y MielínicosDocumento6 páginasEjercicios Neurotróficos y MielínicosJobito Llacta Runa100% (1)
- Conductividad NerviosaDocumento30 páginasConductividad NerviosaJulsAún no hay calificaciones
- Neurofisiologia Del DolorDocumento21 páginasNeurofisiologia Del DolorCHARLES100% (1)
- Actúa ACh y receptoresDocumento8 páginasActúa ACh y receptoresDiana Liz Gamarra ZeñaAún no hay calificaciones
- Mapas 2Documento16 páginasMapas 2Nayareth Baeza Rodriguez100% (1)
- Wuolah Free TEMA 3 FISIOLOGIAMMMMMMMMDocumento4 páginasWuolah Free TEMA 3 FISIOLOGIAMMMMMMMMRodrigo Vallecillo GrauAún no hay calificaciones
- Tabla ComparativaDocumento1 páginaTabla ComparativaMiLthon CastañedaAún no hay calificaciones
- GabaDocumento2 páginasGabaAlvaro CardenasAún no hay calificaciones
- Transmisión Sináptica Med-2Documento44 páginasTransmisión Sináptica Med-2Alexander VillaAún no hay calificaciones
- b54 - Transducción de SeñalesDocumento1 páginab54 - Transducción de SeñalesSanzAún no hay calificaciones
- SinapsisDocumento8 páginasSinapsisloglady00Aún no hay calificaciones
- Taller en Clase SinapsisDocumento6 páginasTaller en Clase SinapsislauraAún no hay calificaciones
- Psicofarmacologia Resumen Lo Esencial en FarmacologiaDocumento52 páginasPsicofarmacologia Resumen Lo Esencial en FarmacologiaRalfAún no hay calificaciones
- Sinapsis: Parcial de Biología 6°MED Yanina Recalde Dos Santos 2021Documento9 páginasSinapsis: Parcial de Biología 6°MED Yanina Recalde Dos Santos 2021Yanina RecaldeAún no hay calificaciones
- Rivera Pico Arlenne 5to B Capitulo 15Documento5 páginasRivera Pico Arlenne 5to B Capitulo 15arlenneriveraAún no hay calificaciones