También podría gustarte
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Calificación: 4.5 de 5 estrellas4.5/5 (119)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaCalificación: 4.5 de 5 estrellas4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe EverandThe Little Book of Hygge: Danish Secrets to Happy LivingCalificación: 3.5 de 5 estrellas3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryCalificación: 3.5 de 5 estrellas3.5/5 (231)
- Grit: The Power of Passion and PerseveranceDe EverandGrit: The Power of Passion and PerseveranceCalificación: 4 de 5 estrellas4/5 (587)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe EverandNever Split the Difference: Negotiating As If Your Life Depended On ItCalificación: 4.5 de 5 estrellas4.5/5 (838)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeCalificación: 4 de 5 estrellas4/5 (5794)
- Rise of ISIS: A Threat We Can't IgnoreDe EverandRise of ISIS: A Threat We Can't IgnoreCalificación: 3.5 de 5 estrellas3.5/5 (137)
- Team of Rivals: The Political Genius of Abraham LincolnDe EverandTeam of Rivals: The Political Genius of Abraham LincolnCalificación: 4.5 de 5 estrellas4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeDe EverandShoe Dog: A Memoir by the Creator of NikeCalificación: 4.5 de 5 estrellas4.5/5 (537)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyCalificación: 3.5 de 5 estrellas3.5/5 (2219)
- The Emperor of All Maladies: A Biography of CancerDe EverandThe Emperor of All Maladies: A Biography of CancerCalificación: 4.5 de 5 estrellas4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreCalificación: 4 de 5 estrellas4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersCalificación: 4.5 de 5 estrellas4.5/5 (344)
- Her Body and Other Parties: StoriesDe EverandHer Body and Other Parties: StoriesCalificación: 4 de 5 estrellas4/5 (821)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceCalificación: 4 de 5 estrellas4/5 (894)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureCalificación: 4.5 de 5 estrellas4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaDe EverandThe Unwinding: An Inner History of the New AmericaCalificación: 4 de 5 estrellas4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)De EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Calificación: 4 de 5 estrellas4/5 (98)
- The Perks of Being a WallflowerDe EverandThe Perks of Being a WallflowerCalificación: 4.5 de 5 estrellas4.5/5 (2099)
- On Fire: The (Burning) Case for a Green New DealDe EverandOn Fire: The (Burning) Case for a Green New DealCalificación: 4 de 5 estrellas4/5 (73)
- VajroliDocumento3 páginasVajroliJithu BayiAún no hay calificaciones
- Important Tips On The Law On SalesDocumento34 páginasImportant Tips On The Law On SalesJanetGraceDalisayFabreroAún no hay calificaciones
- Christoffel Symbols: PHYS 471: Introduction To Relativity and CosmologyDocumento9 páginasChristoffel Symbols: PHYS 471: Introduction To Relativity and Cosmologyarileo3100% (1)
- Bio 140 Exam 4 ReviewDocumento7 páginasBio 140 Exam 4 Reviewlp_blackoutAún no hay calificaciones
- Health Grade 9 1st QuarterDocumento1 páginaHealth Grade 9 1st QuarterCapila Ron100% (1)
- Epd-Lca Meranti Plywood-Final ReportDocumento29 páginasEpd-Lca Meranti Plywood-Final ReporttangAún no hay calificaciones
- Abbas, Al-Arini, Meshikhes - 2014 - Gastric Autonomic Nerve Tumour A Rare Gastric Tumour PDFDocumento2 páginasAbbas, Al-Arini, Meshikhes - 2014 - Gastric Autonomic Nerve Tumour A Rare Gastric Tumour PDFflashjetAún no hay calificaciones
- Abbas Et Al. - 2020 - Maneuvering The Management of A Rare Case of Primary Undifferentiated Cardiac Sarcoma PDFDocumento5 páginasAbbas Et Al. - 2020 - Maneuvering The Management of A Rare Case of Primary Undifferentiated Cardiac Sarcoma PDFflashjetAún no hay calificaciones
- Nana Et Al. - 2014 - Left Renal Vein Thrombosis A Rare Cause of Acute Scrotal Pain PDFDocumento2 páginasNana Et Al. - 2014 - Left Renal Vein Thrombosis A Rare Cause of Acute Scrotal Pain PDFflashjetAún no hay calificaciones
- Nascimento Et Al. - 2013 - Wearable Cardioverter Defibrillator in Stress Cardiomyopathy and Cardiac ArrestDocumento4 páginasNascimento Et Al. - 2013 - Wearable Cardioverter Defibrillator in Stress Cardiomyopathy and Cardiac ArrestflashjetAún no hay calificaciones
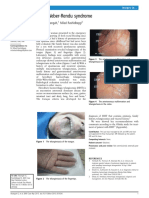
- Abangah, Rashidbeygi - 2013 - Osler-Weber-Rendu SyndromeDocumento2 páginasAbangah, Rashidbeygi - 2013 - Osler-Weber-Rendu SyndromeflashjetAún no hay calificaciones
- Nascimento Et Al. - 2012 - Bloody Nipple Discharge in Infancy - Report of Two CasesDocumento3 páginasNascimento Et Al. - 2012 - Bloody Nipple Discharge in Infancy - Report of Two CasesflashjetAún no hay calificaciones
- Abbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewDocumento5 páginasAbbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewflashjetAún no hay calificaciones
- Abbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewDocumento5 páginasAbbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewflashjetAún no hay calificaciones
- Abbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientDocumento4 páginasAbbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientflashjetAún no hay calificaciones
- Abbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientDocumento4 páginasAbbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientflashjetAún no hay calificaciones
- Namburar Et Al. - 2018 - Waste Generated During Glaucoma Surgery A Comparison of Two Global Facilities PDFDocumento4 páginasNamburar Et Al. - 2018 - Waste Generated During Glaucoma Surgery A Comparison of Two Global Facilities PDFflashjetAún no hay calificaciones
- Nampei Et Al. - 2020 - Comparison of Ocular Surface Squamous Neoplasia and Pterygium Using Anterior Segment Optical Coherence Tomography PDFDocumento4 páginasNampei Et Al. - 2020 - Comparison of Ocular Surface Squamous Neoplasia and Pterygium Using Anterior Segment Optical Coherence Tomography PDFflashjetAún no hay calificaciones
- Nascimento Et Al. - 2012 - Adrenoleukodystrophy A Forgotten Diagnosis in Children With Primary Addison' S DiseaseDocumento5 páginasNascimento Et Al. - 2012 - Adrenoleukodystrophy A Forgotten Diagnosis in Children With Primary Addison' S DiseaseflashjetAún no hay calificaciones
- Nascimento Et Al. - 2013 - Asymptomatic Anomalous Left Anterior Descending Artery Arising From The Right Coronary Artery With A Rare AntDocumento3 páginasNascimento Et Al. - 2013 - Asymptomatic Anomalous Left Anterior Descending Artery Arising From The Right Coronary Artery With A Rare AntflashjetAún no hay calificaciones
- Nassar, Abdul-Jawad, Barnes - 2018 - Hepatic Failure Due To Cholestatic Hepatitis C in An Immunosuppressed Patient Treated With ElbasvirDocumento3 páginasNassar, Abdul-Jawad, Barnes - 2018 - Hepatic Failure Due To Cholestatic Hepatitis C in An Immunosuppressed Patient Treated With ElbasvirflashjetAún no hay calificaciones
- Nasser Et Al. - 2013 - Use of Transoesophageal Echocardiography in Endovascular Stenting For Superior Vena Cava SyndromeDocumento3 páginasNasser Et Al. - 2013 - Use of Transoesophageal Echocardiography in Endovascular Stenting For Superior Vena Cava SyndromeflashjetAún no hay calificaciones
- Nassani Et Al. - 2017 - Foreign Body Penetration Through Jejunal Loops Causing Renal Artery Thrombosis and Renal InfarctDocumento3 páginasNassani Et Al. - 2017 - Foreign Body Penetration Through Jejunal Loops Causing Renal Artery Thrombosis and Renal InfarctflashjetAún no hay calificaciones
- Narula, Nangia - 2014 - Use of An Oscillatory PEP Device To Enhance Bronchial Hygiene in A Patient of post-H1NI Pneumonia and Acute Resp PDFDocumento3 páginasNarula, Nangia - 2014 - Use of An Oscillatory PEP Device To Enhance Bronchial Hygiene in A Patient of post-H1NI Pneumonia and Acute Resp PDFflashjetAún no hay calificaciones
- Nasrat, Al-Alwan - 2012 - Medical Intervention in A Constitutionally Tall ChildDocumento3 páginasNasrat, Al-Alwan - 2012 - Medical Intervention in A Constitutionally Tall ChildflashjetAún no hay calificaciones
- Natrella Et Al. - 2012 - Rare Disease Supratentorial Neurenteric Cyst Associated With A Intraparenchymal SubependymomaDocumento6 páginasNatrella Et Al. - 2012 - Rare Disease Supratentorial Neurenteric Cyst Associated With A Intraparenchymal SubependymomaflashjetAún no hay calificaciones
- Naveen Et Al. - 2014 - Alternatives For Restoration of A Hemisected Mandibular MolarDocumento4 páginasNaveen Et Al. - 2014 - Alternatives For Restoration of A Hemisected Mandibular MolarflashjetAún no hay calificaciones
- Stroke and Transient Ischaemic Attack in Over 16s Diagnosis and Initial Management PDF 975574676437Documento29 páginasStroke and Transient Ischaemic Attack in Over 16s Diagnosis and Initial Management PDF 975574676437MaryamAún no hay calificaciones
- Idiopathic Stroke After Syndromic and Neuromuscular Scoliosis Surgery: A Case Report and Literature ReviewDocumento7 páginasIdiopathic Stroke After Syndromic and Neuromuscular Scoliosis Surgery: A Case Report and Literature ReviewflashjetAún no hay calificaciones
- Nathwani - 2013 - A Complicated Case of AnticoagulationDocumento2 páginasNathwani - 2013 - A Complicated Case of AnticoagulationflashjetAún no hay calificaciones
- Naveen Et Al. - 2013 - Complete Denture With Pharyngeal Bulb ProsthesisDocumento4 páginasNaveen Et Al. - 2013 - Complete Denture With Pharyngeal Bulb ProsthesisflashjetAún no hay calificaciones
- Notes 190915 213814 Ea7 PDFDocumento1 páginaNotes 190915 213814 Ea7 PDFflashjetAún no hay calificaciones
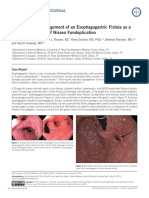
- Naveed Et Al. - 2015 - Diagnosis and Management of An Esophagogastric Fistula As A Rare Complication of Nissen FundoplicationDocumento2 páginasNaveed Et Al. - 2015 - Diagnosis and Management of An Esophagogastric Fistula As A Rare Complication of Nissen FundoplicationflashjetAún no hay calificaciones
- Nguyen Et Al. - 2020 - Human Leukocyte Antigen Susceptibility Map For Severe Acute Respiratory Syndrome Coronavirus 2Documento36 páginasNguyen Et Al. - 2020 - Human Leukocyte Antigen Susceptibility Map For Severe Acute Respiratory Syndrome Coronavirus 2flashjetAún no hay calificaciones
- Nguyen Et Al. - 2019 - Challenges in Management of Autoimmune Hepatitis With Concurrent Graves ThyrotoxicosisDocumento4 páginasNguyen Et Al. - 2019 - Challenges in Management of Autoimmune Hepatitis With Concurrent Graves ThyrotoxicosisflashjetAún no hay calificaciones
- Notes 200430 195226 0c2 PDFDocumento19 páginasNotes 200430 195226 0c2 PDFflashjetAún no hay calificaciones
- Year 9 Autumn 1 2018Documento55 páginasYear 9 Autumn 1 2018andyedwards73Aún no hay calificaciones
- 58 58 International Marketing Chapter WiseDocumento126 páginas58 58 International Marketing Chapter WiseNitish BhaskarAún no hay calificaciones
- Homework Solutions For Chapter 8Documento6 páginasHomework Solutions For Chapter 8api-234237296Aún no hay calificaciones
- HI6025 Assignment T3, 2016Documento3 páginasHI6025 Assignment T3, 2016Abeer ArifAún no hay calificaciones
- Corruption PDFDocumento11 páginasCorruption PDFkaleemullahAún no hay calificaciones
- One, Two and Three PerspectiveDocumento10 páginasOne, Two and Three PerspectiveNikko Bait-itAún no hay calificaciones
- MCD2040 T2 2019 Exam Content and StructureDocumento18 páginasMCD2040 T2 2019 Exam Content and StructureheyitsmemuahAún no hay calificaciones
- CSE Qualifications and PointersDocumento9 páginasCSE Qualifications and PointersChristopher de LeonAún no hay calificaciones
- Reactions of Hydrocarbons and HaloalkanesDocumento6 páginasReactions of Hydrocarbons and HaloalkanesGwynethAdrienneLeighVillapandoAún no hay calificaciones
- Introduction To Logic Syllogisms-1: Class ExerciseDocumento6 páginasIntroduction To Logic Syllogisms-1: Class ExercisePriyanshu PrakashAún no hay calificaciones
- EDMU 520 Phonics Lesson ObservationDocumento6 páginasEDMU 520 Phonics Lesson ObservationElisa FloresAún no hay calificaciones
- Semaphore Twinsoft Manual PDFDocumento101 páginasSemaphore Twinsoft Manual PDFReza AnantoAún no hay calificaciones
- Holliday - 2006 - Native-Speakerism - ELT JournalDocumento3 páginasHolliday - 2006 - Native-Speakerism - ELT JournalBob HowesAún no hay calificaciones
- B1 Mod 01 MathsDocumento152 páginasB1 Mod 01 MathsTharrmaselan VmanimaranAún no hay calificaciones
- Lecture 22 NDocumento6 páginasLecture 22 Ncau toanAún no hay calificaciones
- Case Study Analysis of Six Sigma Implementation Inservice OrganisationsDocumento30 páginasCase Study Analysis of Six Sigma Implementation Inservice OrganisationsMohammed AwolAún no hay calificaciones
- Rehotrical AnalysisDocumento3 páginasRehotrical AnalysisShahid MumtazAún no hay calificaciones
- English March 3Documento7 páginasEnglish March 3Richard ManongsongAún no hay calificaciones
- Webquest Biotechnology RubricDocumento2 páginasWebquest Biotechnology Rubricapi-556285637Aún no hay calificaciones
- Bobby Joe Public NoticeDocumento3 páginasBobby Joe Public NoticeUpscale International InvestmentsAún no hay calificaciones
- King Rama VDocumento3 páginasKing Rama Vsamuel.soo.en.how2324Aún no hay calificaciones
- Assessing Learning Methods and TestsDocumento2 páginasAssessing Learning Methods and TestsZarah Joyce SegoviaAún no hay calificaciones
- In Sunni IslamDocumento5 páginasIn Sunni IslamMk AkangbeAún no hay calificaciones
- 1 Unpacking The SelfDocumento13 páginas1 Unpacking The SelfJEMABEL SIDAYENAún no hay calificaciones