También podría gustarte
- Abbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewDocumento5 páginasAbbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewflashjetAún no hay calificaciones
- Abbas, Al-Arini, Meshikhes - 2014 - Gastric Autonomic Nerve Tumour A Rare Gastric Tumour PDFDocumento2 páginasAbbas, Al-Arini, Meshikhes - 2014 - Gastric Autonomic Nerve Tumour A Rare Gastric Tumour PDFflashjetAún no hay calificaciones
- Abbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientDocumento4 páginasAbbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientflashjetAún no hay calificaciones
- Abbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewDocumento5 páginasAbbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewflashjetAún no hay calificaciones
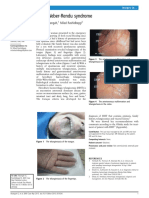
- Abangah, Rashidbeygi - 2013 - Osler-Weber-Rendu SyndromeDocumento2 páginasAbangah, Rashidbeygi - 2013 - Osler-Weber-Rendu SyndromeflashjetAún no hay calificaciones
- Abbas Et Al. - 2020 - Maneuvering The Management of A Rare Case of Primary Undifferentiated Cardiac Sarcoma PDFDocumento5 páginasAbbas Et Al. - 2020 - Maneuvering The Management of A Rare Case of Primary Undifferentiated Cardiac Sarcoma PDFflashjetAún no hay calificaciones
- Nascimento Et Al. - 2012 - Bloody Nipple Discharge in Infancy - Report of Two CasesDocumento3 páginasNascimento Et Al. - 2012 - Bloody Nipple Discharge in Infancy - Report of Two CasesflashjetAún no hay calificaciones
- Nampei Et Al. - 2020 - Comparison of Ocular Surface Squamous Neoplasia and Pterygium Using Anterior Segment Optical Coherence Tomography PDFDocumento4 páginasNampei Et Al. - 2020 - Comparison of Ocular Surface Squamous Neoplasia and Pterygium Using Anterior Segment Optical Coherence Tomography PDFflashjetAún no hay calificaciones
- Abbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientDocumento4 páginasAbbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientflashjetAún no hay calificaciones
- Nascimento Et Al. - 2013 - Wearable Cardioverter Defibrillator in Stress Cardiomyopathy and Cardiac ArrestDocumento4 páginasNascimento Et Al. - 2013 - Wearable Cardioverter Defibrillator in Stress Cardiomyopathy and Cardiac ArrestflashjetAún no hay calificaciones
- Namburar Et Al. - 2018 - Waste Generated During Glaucoma Surgery A Comparison of Two Global Facilities PDFDocumento4 páginasNamburar Et Al. - 2018 - Waste Generated During Glaucoma Surgery A Comparison of Two Global Facilities PDFflashjetAún no hay calificaciones
- Naveen Et Al. - 2013 - Complete Denture With Pharyngeal Bulb ProsthesisDocumento4 páginasNaveen Et Al. - 2013 - Complete Denture With Pharyngeal Bulb ProsthesisflashjetAún no hay calificaciones
- Nana Et Al. - 2014 - Left Renal Vein Thrombosis A Rare Cause of Acute Scrotal Pain PDFDocumento2 páginasNana Et Al. - 2014 - Left Renal Vein Thrombosis A Rare Cause of Acute Scrotal Pain PDFflashjetAún no hay calificaciones
- Nascimento Et Al. - 2013 - Asymptomatic Anomalous Left Anterior Descending Artery Arising From The Right Coronary Artery With A Rare AntDocumento3 páginasNascimento Et Al. - 2013 - Asymptomatic Anomalous Left Anterior Descending Artery Arising From The Right Coronary Artery With A Rare AntflashjetAún no hay calificaciones
- Stroke and Transient Ischaemic Attack in Over 16s Diagnosis and Initial Management PDF 975574676437Documento29 páginasStroke and Transient Ischaemic Attack in Over 16s Diagnosis and Initial Management PDF 975574676437MaryamAún no hay calificaciones
- Narula, Nangia - 2014 - Use of An Oscillatory PEP Device To Enhance Bronchial Hygiene in A Patient of post-H1NI Pneumonia and Acute Resp PDFDocumento3 páginasNarula, Nangia - 2014 - Use of An Oscillatory PEP Device To Enhance Bronchial Hygiene in A Patient of post-H1NI Pneumonia and Acute Resp PDFflashjetAún no hay calificaciones
- Nascimento Et Al. - 2012 - Adrenoleukodystrophy A Forgotten Diagnosis in Children With Primary Addison' S DiseaseDocumento5 páginasNascimento Et Al. - 2012 - Adrenoleukodystrophy A Forgotten Diagnosis in Children With Primary Addison' S DiseaseflashjetAún no hay calificaciones
- Nasrat, Al-Alwan - 2012 - Medical Intervention in A Constitutionally Tall ChildDocumento3 páginasNasrat, Al-Alwan - 2012 - Medical Intervention in A Constitutionally Tall ChildflashjetAún no hay calificaciones
- Nassani Et Al. - 2017 - Foreign Body Penetration Through Jejunal Loops Causing Renal Artery Thrombosis and Renal InfarctDocumento3 páginasNassani Et Al. - 2017 - Foreign Body Penetration Through Jejunal Loops Causing Renal Artery Thrombosis and Renal InfarctflashjetAún no hay calificaciones
- Nassar, Abdul-Jawad, Barnes - 2018 - Hepatic Failure Due To Cholestatic Hepatitis C in An Immunosuppressed Patient Treated With ElbasvirDocumento3 páginasNassar, Abdul-Jawad, Barnes - 2018 - Hepatic Failure Due To Cholestatic Hepatitis C in An Immunosuppressed Patient Treated With ElbasvirflashjetAún no hay calificaciones
- Nathwani - 2013 - A Complicated Case of AnticoagulationDocumento2 páginasNathwani - 2013 - A Complicated Case of AnticoagulationflashjetAún no hay calificaciones
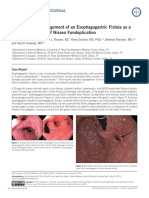
- Naveed Et Al. - 2015 - Diagnosis and Management of An Esophagogastric Fistula As A Rare Complication of Nissen FundoplicationDocumento2 páginasNaveed Et Al. - 2015 - Diagnosis and Management of An Esophagogastric Fistula As A Rare Complication of Nissen FundoplicationflashjetAún no hay calificaciones
- Naveen Et Al. - 2014 - Alternatives For Restoration of A Hemisected Mandibular MolarDocumento4 páginasNaveen Et Al. - 2014 - Alternatives For Restoration of A Hemisected Mandibular MolarflashjetAún no hay calificaciones
- Nasser Et Al. - 2013 - Use of Transoesophageal Echocardiography in Endovascular Stenting For Superior Vena Cava SyndromeDocumento3 páginasNasser Et Al. - 2013 - Use of Transoesophageal Echocardiography in Endovascular Stenting For Superior Vena Cava SyndromeflashjetAún no hay calificaciones
- Natrella Et Al. - 2012 - Rare Disease Supratentorial Neurenteric Cyst Associated With A Intraparenchymal SubependymomaDocumento6 páginasNatrella Et Al. - 2012 - Rare Disease Supratentorial Neurenteric Cyst Associated With A Intraparenchymal SubependymomaflashjetAún no hay calificaciones
- Idiopathic Stroke After Syndromic and Neuromuscular Scoliosis Surgery: A Case Report and Literature ReviewDocumento7 páginasIdiopathic Stroke After Syndromic and Neuromuscular Scoliosis Surgery: A Case Report and Literature ReviewflashjetAún no hay calificaciones
- Nguyen Et Al. - 2020 - Human Leukocyte Antigen Susceptibility Map For Severe Acute Respiratory Syndrome Coronavirus 2Documento36 páginasNguyen Et Al. - 2020 - Human Leukocyte Antigen Susceptibility Map For Severe Acute Respiratory Syndrome Coronavirus 2flashjetAún no hay calificaciones
- Notes 190915 213814 Ea7 PDFDocumento1 páginaNotes 190915 213814 Ea7 PDFflashjetAún no hay calificaciones
- Nguyen Et Al. - 2019 - Challenges in Management of Autoimmune Hepatitis With Concurrent Graves ThyrotoxicosisDocumento4 páginasNguyen Et Al. - 2019 - Challenges in Management of Autoimmune Hepatitis With Concurrent Graves ThyrotoxicosisflashjetAún no hay calificaciones
- Notes 200430 195226 0c2 PDFDocumento19 páginasNotes 200430 195226 0c2 PDFflashjetAún no hay calificaciones
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeCalificación: 4 de 5 estrellas4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeDe EverandShoe Dog: A Memoir by the Creator of NikeCalificación: 4.5 de 5 estrellas4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)De EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Calificación: 4 de 5 estrellas4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceCalificación: 4 de 5 estrellas4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersCalificación: 4.5 de 5 estrellas4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe EverandThe Little Book of Hygge: Danish Secrets to Happy LivingCalificación: 3.5 de 5 estrellas3.5/5 (399)
- Grit: The Power of Passion and PerseveranceDe EverandGrit: The Power of Passion and PerseveranceCalificación: 4 de 5 estrellas4/5 (588)
- The Emperor of All Maladies: A Biography of CancerDe EverandThe Emperor of All Maladies: A Biography of CancerCalificación: 4.5 de 5 estrellas4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaCalificación: 4.5 de 5 estrellas4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe EverandNever Split the Difference: Negotiating As If Your Life Depended On ItCalificación: 4.5 de 5 estrellas4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryCalificación: 3.5 de 5 estrellas3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealDe EverandOn Fire: The (Burning) Case for a Green New DealCalificación: 4 de 5 estrellas4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureCalificación: 4.5 de 5 estrellas4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnDe EverandTeam of Rivals: The Political Genius of Abraham LincolnCalificación: 4.5 de 5 estrellas4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyCalificación: 3.5 de 5 estrellas3.5/5 (2259)
- Rise of ISIS: A Threat We Can't IgnoreDe EverandRise of ISIS: A Threat We Can't IgnoreCalificación: 3.5 de 5 estrellas3.5/5 (137)
- The Unwinding: An Inner History of the New AmericaDe EverandThe Unwinding: An Inner History of the New AmericaCalificación: 4 de 5 estrellas4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreCalificación: 4 de 5 estrellas4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Calificación: 4.5 de 5 estrellas4.5/5 (120)
- The Perks of Being a WallflowerDe EverandThe Perks of Being a WallflowerCalificación: 4.5 de 5 estrellas4.5/5 (2101)
- Her Body and Other Parties: StoriesDe EverandHer Body and Other Parties: StoriesCalificación: 4 de 5 estrellas4/5 (821)
- Osmolarity and IsotonicityDocumento29 páginasOsmolarity and IsotonicityAina'a TammouniAún no hay calificaciones
- 10 Transport in Plants Neet Biology Model Question PaperDocumento5 páginas10 Transport in Plants Neet Biology Model Question PaperJosephAún no hay calificaciones
- Fluid and ElectrolytesDocumento20 páginasFluid and ElectrolytesWyn Agustin100% (1)
- Med Surg Cheat Sheet 1Documento1 páginaMed Surg Cheat Sheet 1Figmentum FigmentumAún no hay calificaciones
- Performance Task 3Documento3 páginasPerformance Task 3Angie CondezaAún no hay calificaciones
- Fluids Concept MappingDocumento1 páginaFluids Concept Mappingmariagarcia415100% (1)
- Different Types of IV FluidsDocumento6 páginasDifferent Types of IV FluidsnicoleAún no hay calificaciones
- Industrial Project Report Magazine For EntrepreneurDocumento17 páginasIndustrial Project Report Magazine For EntrepreneureiribooksAún no hay calificaciones
- Transport Across The Cell Surface MembraneDocumento31 páginasTransport Across The Cell Surface MembraneKalia MckoyAún no hay calificaciones
- CP BIO: Ch. 7 The Cell MembraneDocumento31 páginasCP BIO: Ch. 7 The Cell MembraneJohn Lester BerdinAún no hay calificaciones
- MCQ & Fib-2023Documento26 páginasMCQ & Fib-2023RishiAún no hay calificaciones
- Scheme of Work BIOLOGY FORM 4, 2010Documento28 páginasScheme of Work BIOLOGY FORM 4, 2010WanM.Syamim0% (1)
- Diffusion and Osmosis Worksheet 1Documento2 páginasDiffusion and Osmosis Worksheet 1yojhanvargasAún no hay calificaciones
- AP Biology Lab Review 1-4Documento59 páginasAP Biology Lab Review 1-4swagsurferAún no hay calificaciones
- Notes-Transport in PlantsDocumento15 páginasNotes-Transport in PlantsKisna guptaAún no hay calificaciones
- Pharmaceutical Technology 3 Phar306 Exp 1Documento20 páginasPharmaceutical Technology 3 Phar306 Exp 1zeynep özdalAún no hay calificaciones
- Cell Structure NotesDocumento5 páginasCell Structure NotesJessa Mae LangcuyanAún no hay calificaciones
- 5 HDocumento4 páginas5 HDee GeeAún no hay calificaciones
- OSMOSISDocumento2 páginasOSMOSISHarriet Lucille VidalAún no hay calificaciones
- Saudi Prometric Latest Questions AndanswerDocumento67 páginasSaudi Prometric Latest Questions AndanswerGloria Jaison100% (5)
- CELL TRANSPORTPOWEPOINT (Recovered)Documento140 páginasCELL TRANSPORTPOWEPOINT (Recovered)ntakirutimana valensAún no hay calificaciones
- IV TherapyDocumento34 páginasIV Therapyfairwoods100% (2)
- Introduction To Biology ReviewerDocumento22 páginasIntroduction To Biology ReviewerAnton Miguel Jordan83% (6)
- Suspensions, Colloids and SolutionsDocumento2 páginasSuspensions, Colloids and SolutionsJim Goetz88% (8)
- ScienceDocumento103 páginasScienceWisdom AbotsiAún no hay calificaciones
- Osmolarity LabDocumento2 páginasOsmolarity LabPhi KhanhAún no hay calificaciones
- Biology Grade 10Documento94 páginasBiology Grade 10Ricky NgulubeAún no hay calificaciones
- Hypertonic SolutionsDocumento4 páginasHypertonic SolutionsVanessa PaguiriganAún no hay calificaciones
- Sample Lab Report - Potato OsmosisDocumento6 páginasSample Lab Report - Potato OsmosisCarla Reyes80% (5)
- Resealed ErythrocytesDocumento50 páginasResealed ErythrocytesKrishnareddy Dv100% (1)