También podría gustarte
- Reconocimiento de Celula Animal y VegetalDocumento3 páginasReconocimiento de Celula Animal y Vegetalpedro salvadorAún no hay calificaciones
- Informe Ciclo CelularDocumento3 páginasInforme Ciclo CelularKamilo Valera MorenoAún no hay calificaciones
- Informe de Laboratorio - Reino FungiDocumento16 páginasInforme de Laboratorio - Reino FungiGabriel Calvo MolinaAún no hay calificaciones
- Reconocimiento Células SanguíneasDocumento5 páginasReconocimiento Células SanguíneasLucia Arriaga DAún no hay calificaciones
- Actividad 06 Biologia - El Ciclo Celular Eucariote y El CancerDocumento5 páginasActividad 06 Biologia - El Ciclo Celular Eucariote y El CancerCarlos Damian ParedesAún no hay calificaciones
- Practica CiclosisDocumento5 páginasPractica CiclosisJhohan Andersson Sanchez YlquimicheAún no hay calificaciones
- Introducción de La MitosisDocumento2 páginasIntroducción de La MitosisJuan Espinoza CcoraAún no hay calificaciones
- Importancia Del Tamaño CelularDocumento2 páginasImportancia Del Tamaño CelularAlfonso Rada Vargas50% (2)
- Clase 4 TrofoblastoDocumento21 páginasClase 4 TrofoblastoPriscilla Sandoya BernabéAún no hay calificaciones
- CONCLUSIÓNDocumento1 páginaCONCLUSIÓNNatán MarcilloAún no hay calificaciones
- Trabajo de Biologia 2 TrimestreDocumento14 páginasTrabajo de Biologia 2 TrimestreAnaGabrielaPérezAún no hay calificaciones
- Laboratorio Difusion, Osmosis y DialisisDocumento11 páginasLaboratorio Difusion, Osmosis y DialisisMauricio Cedeño36% (14)
- Informe Fenómenos Físicos de La CelulaDocumento17 páginasInforme Fenómenos Físicos de La CelulaFacultad Medicina SJBAún no hay calificaciones
- Observacion de Celulas Sanguineas #6Documento11 páginasObservacion de Celulas Sanguineas #6Felipe Gordillo100% (1)
- Formas de núcleo en células sanguíneasDocumento4 páginasFormas de núcleo en células sanguíneasDaniel Marquiño Alonzo Paz50% (2)
- CentriolosDocumento2 páginasCentriolosPaola AyalaAún no hay calificaciones
- Normas Bioseguridad Lab QuímicaDocumento15 páginasNormas Bioseguridad Lab QuímicaJohn Hector Lopez Gomez100% (1)
- Enfermedades Citoesqueléticas y de OrganelosDocumento5 páginasEnfermedades Citoesqueléticas y de OrganelosPedro Dominguez50% (2)
- Informe #3 A. Clinicos LeucocitosDocumento6 páginasInforme #3 A. Clinicos Leucocitos02joseluis02Aún no hay calificaciones
- Monografia de Histologia Tejido SanguineoDocumento25 páginasMonografia de Histologia Tejido SanguineoTener Papagayo0% (2)
- Estudio proteínas enzimas enfermeríaDocumento1 páginaEstudio proteínas enzimas enfermeríaVivian GarayAún no hay calificaciones
- Embriologia Del Corazon PDFDocumento5 páginasEmbriologia Del Corazon PDFRuben Dario SierraAún no hay calificaciones
- DGB y DGT PresentaciónDocumento36 páginasDGB y DGT PresentaciónEdgaris Sc'r0% (1)
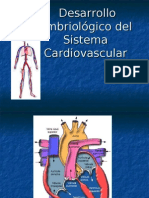
- Desarrollo Embriologico Del Sistema CardiovascularDocumento46 páginasDesarrollo Embriologico Del Sistema CardiovascularKevin Flores100% (4)
- Marco Teorico Sobre LeishmaniasisDocumento8 páginasMarco Teorico Sobre LeishmaniasisSG SimónAún no hay calificaciones
- Informe de LaboratorioDocumento6 páginasInforme de LaboratorioERIKA RUIZAún no hay calificaciones
- Enlace Que Se Forma en La Molécula de HNO3 Mediante El Punto Electrón de LewisDocumento6 páginasEnlace Que Se Forma en La Molécula de HNO3 Mediante El Punto Electrón de LewisSydney FernandezAún no hay calificaciones
- Osmosis Del HuevoDocumento10 páginasOsmosis Del HuevoYoselyn Jhoana Limache Marin0% (1)
- Discusion MeiosisDocumento1 páginaDiscusion MeiosisJosesithaRodriguezLopezAún no hay calificaciones
- Transporte celular a través de membranas: ósmosis, difusión, bombas iónicasDocumento9 páginasTransporte celular a través de membranas: ósmosis, difusión, bombas iónicasPipe Kllejas0% (1)
- Observación de Protozoarios Estudiados en ClaseDocumento14 páginasObservación de Protozoarios Estudiados en ClaseCésar ArteagaAún no hay calificaciones
- Biofisica de La Respiracion...Documento16 páginasBiofisica de La Respiracion...Anonymous h68MYzn2INAún no hay calificaciones
- Histología de Órganos LinfoidesDocumento14 páginasHistología de Órganos LinfoidesRosyAún no hay calificaciones
- Ciclo de la comunicación y sus elementosDocumento12 páginasCiclo de la comunicación y sus elementosktolkrAún no hay calificaciones
- Solicitud Unl FebreroDocumento1 páginaSolicitud Unl FebreroAleJiménezAún no hay calificaciones
- Ciclo de Vida de La Taenia SoliumDocumento5 páginasCiclo de Vida de La Taenia SoliumJuan Abel CristencensAún no hay calificaciones
- Informe GyE UPAO Semana 6Documento13 páginasInforme GyE UPAO Semana 6Alejandra Quino HerreraAún no hay calificaciones
- Mapas ConceptualesDocumento10 páginasMapas ConceptualesDany CancheAún no hay calificaciones
- Identificacion de Grupos Funcionales FinalDocumento11 páginasIdentificacion de Grupos Funcionales FinalLuis Enrique Zamora40% (5)
- Introducción a la MicologíaDocumento11 páginasIntroducción a la MicologíaomelisAún no hay calificaciones
- Permeabilidad LaboratorioDocumento11 páginasPermeabilidad LaboratoriomelyAún no hay calificaciones
- Resultados de Cromatina Sexual en Mujeres y VaronesDocumento4 páginasResultados de Cromatina Sexual en Mujeres y VaronesdelfinaAún no hay calificaciones
- Farmacoterapia en El EmbarazoDocumento20 páginasFarmacoterapia en El EmbarazoInternational Medical Publisher100% (1)
- Informe Mitosis 1Documento7 páginasInforme Mitosis 1Constantine1127100% (1)
- Informe ElectrolitosDocumento13 páginasInforme Electrolitosjuandgh33% (3)
- Soluciones en El Cuerpo HumanoDocumento27 páginasSoluciones en El Cuerpo HumanoGiancarlo David Viviano Reyes0% (3)
- Adme de AlcoholDocumento24 páginasAdme de AlcoholAnonymous Wwec0BEPJmAún no hay calificaciones
- Referencia Al Csmi El BosqueDocumento5 páginasReferencia Al Csmi El BosqueAlois Silva UgazAún no hay calificaciones
- OvogénesisDocumento5 páginasOvogénesisKevin CisAún no hay calificaciones
- Unidad y Diversidad de Las CélulasDocumento6 páginasUnidad y Diversidad de Las CélulasRita Maribel Bonilla Mora0% (1)
- Caso Clínico Semana 1Documento36 páginasCaso Clínico Semana 1Mirella Ojeda FloresAún no hay calificaciones
- Musculo Liso 2Documento16 páginasMusculo Liso 2Armando ponce100% (1)
- Resumen 1 Capítulo de Introducción A La BotánicaDocumento5 páginasResumen 1 Capítulo de Introducción A La BotánicaJackeline Marin100% (1)
- Analisis de LA CÉLULADocumento3 páginasAnalisis de LA CÉLULAAyleneAún no hay calificaciones
- Ensayo de Patologia o PatologiasDocumento12 páginasEnsayo de Patologia o Patologiasyiyo al daoud50% (2)
- Taller Laboratorio de BiologíaDocumento4 páginasTaller Laboratorio de BiologíaElizabeth FerrerAún no hay calificaciones
- Fundamento TeóricoDocumento2 páginasFundamento Teóricololys12380% (10)
- Celulas EucariotasDocumento17 páginasCelulas EucariotasUSMP FN ARCHIVOSAún no hay calificaciones
- Informe Mitosis en Celula VegetalDocumento11 páginasInforme Mitosis en Celula VegetalJaviera San MartinAún no hay calificaciones
- Ciclo celular y mitosisDocumento6 páginasCiclo celular y mitosisMaría Laura Rodríguez SayagoAún no hay calificaciones
- Rhe 10404452679 e 00126Documento1 páginaRhe 10404452679 e 00126wilmerAún no hay calificaciones
- ALOPURINOL1Documento14 páginasALOPURINOL1wilmerAún no hay calificaciones
- Acta Del Comité de Defensa de Los Intereses de La Cooperativa AgrariaDocumento3 páginasActa Del Comité de Defensa de Los Intereses de La Cooperativa AgrariawilmerAún no hay calificaciones
- Rhe10404452679e00126 PDFDocumento1 páginaRhe10404452679e00126 PDFwilmerAún no hay calificaciones
- Triptico Con Diseño Muy Bueno Hecho Por Rosa María PDFDocumento2 páginasTriptico Con Diseño Muy Bueno Hecho Por Rosa María PDFwilmerAún no hay calificaciones
- Abrahan Valdelomar1Documento23 páginasAbrahan Valdelomar1wilmer81280Aún no hay calificaciones
- Monografia Uña de GatoDocumento31 páginasMonografia Uña de Gatowilmer81280100% (2)
- Contrato de Alquiler de LocalDocumento3 páginasContrato de Alquiler de LocalwilmerAún no hay calificaciones
- Monografia Uña de GatoDocumento31 páginasMonografia Uña de Gatowilmer81280100% (2)
- El impacto de los correos electrónicosDocumento24 páginasEl impacto de los correos electrónicoswilmer8128040% (5)
- Crecimiento de cabello con Coca ColaDocumento8 páginasCrecimiento de cabello con Coca ColawilmerAún no hay calificaciones
- Trabajo de Investigacion de Insecticida Cacera de AjosDocumento13 páginasTrabajo de Investigacion de Insecticida Cacera de AjoswilmerAún no hay calificaciones
- Tema 1 Una Función Química La Vida - 2021Documento37 páginasTema 1 Una Función Química La Vida - 2021NataliaAún no hay calificaciones
- Tarea 0 - Reconocer La MicrobiologiaDocumento15 páginasTarea 0 - Reconocer La Microbiologiayesica lorena villa blandonAún no hay calificaciones
- PROTOCOLODELABORATORIOBIOLOGIACELULARDocumento140 páginasPROTOCOLODELABORATORIOBIOLOGIACELULARCamilo Caicedo0% (1)
- Técnicas HistológicasDocumento13 páginasTécnicas HistológicasFeddAún no hay calificaciones
- Las BacteriasDocumento9 páginasLas BacteriasDimas Rangel Guerrero RamirezAún no hay calificaciones
- Clase 1. Aplicaciones de La GeneÌ Tica y BiologiÌ A MolecularDocumento31 páginasClase 1. Aplicaciones de La GeneÌ Tica y BiologiÌ A MolecularfridsaAún no hay calificaciones
- TEMA 1 Microbiologia de Los AlimentosDocumento47 páginasTEMA 1 Microbiologia de Los AlimentosValeria Bianca Martínez ParcoAún no hay calificaciones
- Syllabus Del Curso Biología Celular y MolecularDocumento10 páginasSyllabus Del Curso Biología Celular y MolecularpeterordosgoitiapeteAún no hay calificaciones
- Guía para El Desarrollo Del Componente Práctico y Rúbrica de Evaluación - Unidades 1 y 2 - Tarea 4 - Asistir Al Laboratorio de BiologíaDocumento10 páginasGuía para El Desarrollo Del Componente Práctico y Rúbrica de Evaluación - Unidades 1 y 2 - Tarea 4 - Asistir Al Laboratorio de BiologíaSergio Luis Pacheco ColonAún no hay calificaciones
- Cien y Éti p3t01 PDFDocumento4 páginasCien y Éti p3t01 PDFharol stiven martinezAún no hay calificaciones
- Bioquimica y Biologia MolecularDocumento259 páginasBioquimica y Biologia MolecularJesusChumbipuma100% (1)
- Agnes Pockels - Revista Contactos 87Documento62 páginasAgnes Pockels - Revista Contactos 87Christina MillerAún no hay calificaciones
- 1 ESO - U7 - Actividades AmpliaciónDocumento2 páginas1 ESO - U7 - Actividades Ampliaciónalejandrosantorcaz97Aún no hay calificaciones
- Guia Biologia Ii-Prueba 8-LDDocumento362 páginasGuia Biologia Ii-Prueba 8-LDyerko corderoAún no hay calificaciones
- Ejemplos de Textos Expositivos y Teoría de Textos Expositivos y Argumentativos.Documento16 páginasEjemplos de Textos Expositivos y Teoría de Textos Expositivos y Argumentativos.All Joao Paredes Quispe100% (1)
- Clase 2 - La Celula, Tipos, Funciones - 2023-IDocumento44 páginasClase 2 - La Celula, Tipos, Funciones - 2023-IJIMENA MAGALY JARAMILLO CORDOVAAún no hay calificaciones
- Formato A - Lab Microbiología 1Documento5 páginasFormato A - Lab Microbiología 1Juan Alberto Rimarachin AltamiranoAún no hay calificaciones
- Programa 52223 Microbiología GeneralDocumento13 páginasPrograma 52223 Microbiología GeneralChristian ArevaloAún no hay calificaciones
- HongosDocumento7 páginasHongosPaola VillanuevaAún no hay calificaciones
- Monografia Templo: Orden RosacruzDocumento7 páginasMonografia Templo: Orden RosacruzCarlos Alberto Casado100% (1)
- Histología-Guia - Practica 02Documento10 páginasHistología-Guia - Practica 02Eliane Marreros BazauriAún no hay calificaciones
- Folleto Infantil para Actividades Proyectos y Juegos Con Niños Doodle Dibujado Turquesa y RojoDocumento2 páginasFolleto Infantil para Actividades Proyectos y Juegos Con Niños Doodle Dibujado Turquesa y RojoMavi MagdielAún no hay calificaciones
- Sexto PrimariaDocumento5 páginasSexto Primariaperry_moAún no hay calificaciones
- Genoma HumanoDocumento2 páginasGenoma HumanoJuan Lopez LopezAún no hay calificaciones
- Enstc414 A23v2Documento52 páginasEnstc414 A23v2Oliver MorosoAún no hay calificaciones
- Mecanismos de Proteccion de La NitrogenasaDocumento12 páginasMecanismos de Proteccion de La NitrogenasaKaren AlejandreAún no hay calificaciones
- Entrevista Desde El Enfoque BiodescodificadorDocumento6 páginasEntrevista Desde El Enfoque BiodescodificadorVeronica Cruz GuillenAún no hay calificaciones
- Practica de Laboratorio Acidos Nucleicos y NucleoDocumento11 páginasPractica de Laboratorio Acidos Nucleicos y NucleoCarlOz ContrerasAún no hay calificaciones
- Cuadernillo de Ingreso 2020 2do AñoDocumento67 páginasCuadernillo de Ingreso 2020 2do AñoHectorAún no hay calificaciones
- Escalante Diana - IP MEMBRANAS PDFDocumento10 páginasEscalante Diana - IP MEMBRANAS PDFDiana EscalanteAún no hay calificaciones