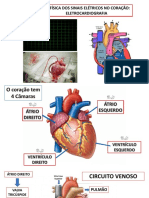
También podría gustarte
- Tabela CarrapatoDocumento1 páginaTabela CarrapatoDeyseRibeiroAún no hay calificaciones
- Farmacologia e FarmacocinéticaDocumento12 páginasFarmacologia e FarmacocinéticaeduardaAún no hay calificaciones
- Mapa Mental - GENERO DEMODEXDocumento4 páginasMapa Mental - GENERO DEMODEXGabriella do CarmoAún no hay calificaciones
- Sistema ImunitárioDocumento10 páginasSistema ImunitárioMargarida FonsecaAún no hay calificaciones
- Apostila de Patologia GeralDocumento81 páginasApostila de Patologia Geralguilherme ribeiroAún no hay calificaciones
- BabesioseDocumento29 páginasBabesioseVictoria RomanoAún no hay calificaciones
- Tecido Muscular..Documento8 páginasTecido Muscular..GARDON SINVALDOAún no hay calificaciones
- Apostilinha IMUNODocumento109 páginasApostilinha IMUNOIngridAún no hay calificaciones
- Apostila Resumo Fisiologia AnimalDocumento68 páginasApostila Resumo Fisiologia AnimalNatália Ruiz Heiderich RozaAún no hay calificaciones
- Teniase e CisticercoseDocumento28 páginasTeniase e CisticercoseHelirius DeitosAún no hay calificaciones
- 2 Imunidade InataDocumento14 páginas2 Imunidade InataLeticia Costa100% (1)
- Biologia - DOMÍNIO ARCHAEADocumento16 páginasBiologia - DOMÍNIO ARCHAEArafael silvaAún no hay calificaciones
- Teniase e Cisticercose PDFDocumento20 páginasTeniase e Cisticercose PDFJeremy S. FölderAún no hay calificaciones
- 1 HematologiaDocumento41 páginas1 HematologiaAna Cláudia SouzaAún no hay calificaciones
- Fisiologia Do Sistema RenalDocumento17 páginasFisiologia Do Sistema RenalFernanda AmaralAún no hay calificaciones
- Medicina Veterinária Investigativa (Patologia Veterinária) - ResumoDocumento9 páginasMedicina Veterinária Investigativa (Patologia Veterinária) - ResumoKauã SantosAún no hay calificaciones
- Coleta, Armazenamento e Envio de Amostras de Material de Aborto para Diagnóstico de Doenças Infecciosas em BovinosDocumento6 páginasColeta, Armazenamento e Envio de Amostras de Material de Aborto para Diagnóstico de Doenças Infecciosas em BovinosjaimecafigueroaAún no hay calificaciones
- Hidatidose e Himenolepiase - P2 ParasitoDocumento2 páginasHidatidose e Himenolepiase - P2 Parasitovitoryagomes02Aún no hay calificaciones
- HemoparasitosDocumento17 páginasHemoparasitosFrancieli Malvestio BlaserAún no hay calificaciones
- Hemoparasitose - Aula 7Documento12 páginasHemoparasitose - Aula 7jessicaAún no hay calificaciones
- Doenças Infecciosas GrandesDocumento78 páginasDoenças Infecciosas GrandesALISON100% (1)
- Introdução Semiologia 22-23Documento21 páginasIntrodução Semiologia 22-23Andressa SouzaAún no hay calificaciones
- Apostila ParasitoDocumento274 páginasApostila ParasitoJaneteElenice50% (2)
- Propriedade Das Respostas ImunesDocumento5 páginasPropriedade Das Respostas ImunesLara BorgesAún no hay calificaciones
- Morte CelularDocumento34 páginasMorte CelularEdson LuccasAún no hay calificaciones
- Adenite Equina - GarrotilhoDocumento26 páginasAdenite Equina - Garrotilhobhanny2Aún no hay calificaciones
- Helmintologia Veterinária - Introdução - Aula 1Documento33 páginasHelmintologia Veterinária - Introdução - Aula 1Aline KoslinskiAún no hay calificaciones
- Genética de PopulaçõesDocumento47 páginasGenética de PopulaçõesAmanda CristinaAún no hay calificaciones
- P1Donçasicos: Aula 2 - ColibaciloseDocumento17 páginasP1Donçasicos: Aula 2 - ColibaciloseNicolle de Alcantara Zuchini Nogueira SoaresAún no hay calificaciones
- Patologia VeterináriaDocumento11 páginasPatologia Veterináriagabrielly PadilhaAún no hay calificaciones
- Patologia VeterinariaDocumento14 páginasPatologia VeterinariabiancapaluAún no hay calificaciones
- 04 MiologiaDocumento34 páginas04 MiologiaThais Nunes100% (1)
- Coccidiose Eimeriose e CriotosporidioseDocumento6 páginasCoccidiose Eimeriose e CriotosporidioseAna Rita VelhoAún no hay calificaciones
- Fungos - Introdução VeterináriaDocumento18 páginasFungos - Introdução VeterináriaDalila CaixetaAún no hay calificaciones
- Sistema CardiovascularDocumento54 páginasSistema CardiovascularDeboraAún no hay calificaciones
- Pulgas Piolhos e BarbeirosDocumento88 páginasPulgas Piolhos e BarbeirosJoao Vitor Pereira Marques BorgesAún no hay calificaciones
- Estudodirigido OsteologiaDocumento2 páginasEstudodirigido OsteologiaAnnyelle BeatrizAún no hay calificaciones
- Micologia Aula 1Documento48 páginasMicologia Aula 1Walter MoreiraAún no hay calificaciones
- 2-Eimeria e IsosporaDocumento72 páginas2-Eimeria e IsosporaCarolina Silva100% (3)
- Medicina Veterinária - Patologia Sistema UrinárioDocumento5 páginasMedicina Veterinária - Patologia Sistema UrinárioJulia Bicalho100% (1)
- Estudo Dirigido ParasitologiaDocumento11 páginasEstudo Dirigido ParasitologiaThays Couto BarbosaAún no hay calificaciones
- CLÍNICA DE SELVAGENS Resumo Prova 1Documento38 páginasCLÍNICA DE SELVAGENS Resumo Prova 1Marcelle Ferreira100% (1)
- Patologia - Modulo 4 2022Documento23 páginasPatologia - Modulo 4 2022Ana Carolina da SilvaAún no hay calificaciones
- Resumo Histologia Do Sistema CardiovascularDocumento8 páginasResumo Histologia Do Sistema CardiovascularCleyvison MonteiroAún no hay calificaciones
- Carrapatos e Arcaros (Teórica e Prática) - Parasitologia VeterináriaDocumento36 páginasCarrapatos e Arcaros (Teórica e Prática) - Parasitologia Veterináriarafaela souzaAún no hay calificaciones
- Ciclo Do Trypanosoma CruziDocumento7 páginasCiclo Do Trypanosoma CruziNathália SilvaAún no hay calificaciones
- Aula 4 - Vias de AdministraçãoDocumento30 páginasAula 4 - Vias de AdministraçãoAlvaro GaldosAún no hay calificaciones
- ED DeontologiaDocumento4 páginasED Deontologiascbecker10006228Aún no hay calificaciones
- Apostila Farmacologia 5periodoDocumento50 páginasApostila Farmacologia 5periodoLetycia AmorinAún no hay calificaciones
- Sistema Cardiovascular - Passei DiretoDocumento6 páginasSistema Cardiovascular - Passei Diretotata m.l.Aún no hay calificaciones
- Ciclo de Krebs e Fosforilação OxidativaDocumento56 páginasCiclo de Krebs e Fosforilação OxidativaMariana Gonçalves MacedoAún no hay calificaciones
- Planos e Eixos Veterinária 1 SemestreDocumento26 páginasPlanos e Eixos Veterinária 1 SemestreClaudia SilvaAún no hay calificaciones
- Aula 06 - BabesiidaeDocumento41 páginasAula 06 - BabesiidaeGabrieli MolinaAún no hay calificaciones
- Aula 5-12-04 - 2022 Biofísica - UnipDocumento47 páginasAula 5-12-04 - 2022 Biofísica - Uniplauana RebecaAún no hay calificaciones
- Emergencias Na Anestesia VeterinariaDocumento5 páginasEmergencias Na Anestesia VeterinariaAle AlssufeAún no hay calificaciones
- 607b3d5e93435 Linfocitos-TDocumento15 páginas607b3d5e93435 Linfocitos-TTaís BoaventuraAún no hay calificaciones
- Imunologia 04Documento31 páginasImunologia 04karolineandrade389Aún no hay calificaciones
- IMUNIDADEDocumento4 páginasIMUNIDADERebecca DanneckerAún no hay calificaciones
- 6 Ativação de Linfocito TDocumento11 páginas6 Ativação de Linfocito TLeticia CostaAún no hay calificaciones
- RastreamentoDocumento97 páginasRastreamentoMohema DuarteAún no hay calificaciones
- Ramais Ebserh HC-UFTMDocumento15 páginasRamais Ebserh HC-UFTMthiago_eliseuAún no hay calificaciones
- ED2 (Dor Aguda e Crônica + Fibromialgia)Documento6 páginasED2 (Dor Aguda e Crônica + Fibromialgia)thiago_eliseuAún no hay calificaciones
- 5 - Lesões GenitaisDocumento8 páginas5 - Lesões Genitaisthiago_eliseuAún no hay calificaciones
- Atividade 1 - ED1 (Crendices Populares)Documento11 páginasAtividade 1 - ED1 (Crendices Populares)thiago_eliseuAún no hay calificaciones
- Método CornelDocumento6 páginasMétodo Cornelthiago_eliseuAún no hay calificaciones
- 23229e-DC Manejo Da Febre Aguda SITEDocumento14 páginas23229e-DC Manejo Da Febre Aguda SITEAngelo Bedim FranzoniAún no hay calificaciones
- Ramos-Casals2014 en PTDocumento14 páginasRamos-Casals2014 en PTthiago_eliseuAún no hay calificaciones
- Crendices Populares MarcadoDocumento8 páginasCrendices Populares Marcadothiago_eliseuAún no hay calificaciones
- 5 K VNKNQ JWSBF RT7 BX GDFSJBDocumento3 páginas5 K VNKNQ JWSBF RT7 BX GDFSJBthiago_eliseuAún no hay calificaciones
- BGvyw CTFC RX BS5 FD 9 ZVP 4 Q FDocumento5 páginasBGvyw CTFC RX BS5 FD 9 ZVP 4 Q Fthiago_eliseuAún no hay calificaciones
- PanhipopituitarismoDocumento4 páginasPanhipopituitarismothiago_eliseuAún no hay calificaciones
- Rita TMagalhaesDocumento46 páginasRita TMagalhaesthiago_eliseuAún no hay calificaciones
- 23358c-DC-Pneumonias Adquiridas Nao Complicadas - InddDocumento11 páginas23358c-DC-Pneumonias Adquiridas Nao Complicadas - Inddthiago_eliseuAún no hay calificaciones
- MINIGUIA - Especialidades PDFDocumento36 páginasMINIGUIA - Especialidades PDFLuiz HenriqueAún no hay calificaciones
- Semiologia OrtopedicaDocumento13 páginasSemiologia OrtopedicarenarrudaAún no hay calificaciones
- T01 - Questionário 1 - ReumatoDocumento3 páginasT01 - Questionário 1 - Reumatothiago_eliseuAún no hay calificaciones
- Supra R RenalDocumento6 páginasSupra R Renalthiago_eliseuAún no hay calificaciones
- CASO CLÍNICO 1 - INSUFICIÊNCIA ADRENAL - Thiago Henrique EliseuDocumento1 páginaCASO CLÍNICO 1 - INSUFICIÊNCIA ADRENAL - Thiago Henrique Eliseuthiago_eliseuAún no hay calificaciones
- T01 - Introdução À Reumatologia (Transcrição)Documento3 páginasT01 - Introdução À Reumatologia (Transcrição)thiago_eliseuAún no hay calificaciones
- Educacao Medica Desafio Da Humanizacao Na FormacaoDocumento12 páginasEducacao Medica Desafio Da Humanizacao Na Formacaothiago_eliseuAún no hay calificaciones
- Aula 2 Pessotti PDFDocumento9 páginasAula 2 Pessotti PDFthiago_eliseuAún no hay calificaciones
- Pan HipopituitarismoDocumento5 páginasPan Hipopituitarismothiago_eliseuAún no hay calificaciones
- SBGG Guia de DorDocumento32 páginasSBGG Guia de DorGessyPbAún no hay calificaciones
- Supra R RenalDocumento6 páginasSupra R Renalthiago_eliseuAún no hay calificaciones
- Hipotálamo e HipófiseDocumento9 páginasHipotálamo e Hipófisethiago_eliseuAún no hay calificaciones
- TIREOIDE - ANATOMIA, EMBRIOLOGIA E FISIOLOGIA - 20-08-2019 - TerçaDocumento8 páginasTIREOIDE - ANATOMIA, EMBRIOLOGIA E FISIOLOGIA - 20-08-2019 - Terçathiago_eliseuAún no hay calificaciones
- Insuficiencia Aortica-RespostasDocumento4 páginasInsuficiencia Aortica-Respostasthiago_eliseuAún no hay calificaciones
- EletroDocumento5 páginasEletrothiago_eliseuAún no hay calificaciones
- Curso de Eletrocardiograma - TaquiDocumento12 páginasCurso de Eletrocardiograma - Taquithiago_eliseuAún no hay calificaciones
- Como Se Há de Haver o Senhor - AntonilDocumento2 páginasComo Se Há de Haver o Senhor - AntonilPlinio LabriolaAún no hay calificaciones
- Folder Terraco Urban - Baixa PDFDocumento13 páginasFolder Terraco Urban - Baixa PDFxoxotadayAún no hay calificaciones
- Tecnicas Especiais de InjeçãoDocumento3 páginasTecnicas Especiais de InjeçãoViniciusAún no hay calificaciones
- Aula 1 - Fundações 13-08-2020Documento5 páginasAula 1 - Fundações 13-08-2020José NunesAún no hay calificaciones
- Apostila TEAPDocumento7 páginasApostila TEAPErick Natividad ZevallosAún no hay calificaciones
- Sika Protecção Betão - 03.223Documento3 páginasSika Protecção Betão - 03.223FilipeBaptistaAún no hay calificaciones
- Descobrimento Do Brasil - Cola Da WebDocumento4 páginasDescobrimento Do Brasil - Cola Da WebWallace O RoqueAún no hay calificaciones
- Aval - Diag Mat 4 Ano 2014Documento7 páginasAval - Diag Mat 4 Ano 2014Sidilaine AlvesAún no hay calificaciones
- CLR James - Os Jacobinos Negros-Boitempo (2010)Documento202 páginasCLR James - Os Jacobinos Negros-Boitempo (2010)tadeu100% (6)
- Aula-2-Termodinâmica-e-Equilibrio Experimental - Tensão SuperficialDocumento5 páginasAula-2-Termodinâmica-e-Equilibrio Experimental - Tensão SuperficialEstefani SousaAún no hay calificaciones
- Espaços Motores de Fluxos Mundiais (12.º)Documento10 páginasEspaços Motores de Fluxos Mundiais (12.º)profgeofernando100% (2)
- RelatorioDocumento3 páginasRelatorioLaura Maia AntunesAún no hay calificaciones
- O Liberalismo É PecadoDocumento192 páginasO Liberalismo É PecadoLuiza Colassanto ZamboliAún no hay calificaciones
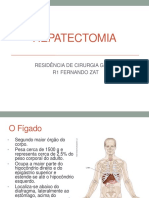
- HEPATECTOMIADocumento34 páginasHEPATECTOMIAFernando ZatAún no hay calificaciones
- Rito Da Dedicação de Uma IgrejaDocumento9 páginasRito Da Dedicação de Uma Igrejastorealcateia22Aún no hay calificaciones
- Tds Ficha - Tecnica Preg - Facil Rev.06 17docDocumento2 páginasTds Ficha - Tecnica Preg - Facil Rev.06 17docMarcosAún no hay calificaciones
- ShabtiDocumento6 páginasShabtinicollas andrewAún no hay calificaciones
- 12 FP MensagemDocumento6 páginas12 FP MensagemAna Francisca MonteiroAún no hay calificaciones
- Lista de Exercícios 1 - Econometria - 2019-2Documento9 páginasLista de Exercícios 1 - Econometria - 2019-2RenanMüllerAún no hay calificaciones
- Atividade Cultura Brasileira 1 TEXTOSDocumento1 páginaAtividade Cultura Brasileira 1 TEXTOSRafael FrancoAún no hay calificaciones
- Apostila Mariologia - Pe. João Paulo VelosoDocumento25 páginasApostila Mariologia - Pe. João Paulo VelosoTarcisio Augusto100% (1)
- Relatório Medidas e IncertezasDocumento9 páginasRelatório Medidas e IncertezasLeonardo A. RuschAún no hay calificaciones
- Portugues 12o Resumos Importantes para o ExameDocumento35 páginasPortugues 12o Resumos Importantes para o ExameManuel SousaAún no hay calificaciones
- Sucesso - Português 4Documento336 páginasSucesso - Português 4Camilla Antunes0% (1)
- Cantos para A Quaresma 2024Documento5 páginasCantos para A Quaresma 2024Keven RogerAún no hay calificaciones
- Lista de Exercícios 9 - Matemática - 4º BimestreDocumento4 páginasLista de Exercícios 9 - Matemática - 4º BimestreLuís Felipe Peterlini75% (4)
- CoagulogramaDocumento27 páginasCoagulogramaJessica Teixeira100% (1)
- Acentuação GráficaDocumento3 páginasAcentuação GráficaAdrianaAún no hay calificaciones
- Tesao Vaca05Documento7 páginasTesao Vaca05dsfsdfsAún no hay calificaciones
- Catalago Guaporé Proteção 2019 1.0 PDFDocumento12 páginasCatalago Guaporé Proteção 2019 1.0 PDFmaylon campystAún no hay calificaciones