También podría gustarte
- Red Cell Membrane and Cation Deficiency in RH Null SyndromeDocumento10 páginasRed Cell Membrane and Cation Deficiency in RH Null SyndromeRoyaAún no hay calificaciones
- 3 - Predicting The Morphology of Sickle Red Blood Cells Using Coarse-GrainedDocumento10 páginas3 - Predicting The Morphology of Sickle Red Blood Cells Using Coarse-GrainedAli M. RiyathAún no hay calificaciones
- Intrinsic RBC DisordersDocumento4 páginasIntrinsic RBC DisordersMartin ClydeAún no hay calificaciones
- Full Dynamics of A Red Blood Cell in Shear Ow: 20808 - 20813 Pnas December 18, 2012 Vol. 109 No. 51Documento6 páginasFull Dynamics of A Red Blood Cell in Shear Ow: 20808 - 20813 Pnas December 18, 2012 Vol. 109 No. 51rezqAún no hay calificaciones
- Abnormal RBC Morphology PMB and MwaDocumento2 páginasAbnormal RBC Morphology PMB and MwaPaul BandaAún no hay calificaciones
- 2 Abo Blood Group SystemDocumento7 páginas2 Abo Blood Group SystempearlsAún no hay calificaciones
- Hema Lec #7 Morphological Evaluation of ErythrocytesDocumento11 páginasHema Lec #7 Morphological Evaluation of ErythrocytesnadeslemonAún no hay calificaciones
- Fatal pulmonary leukostasis following AML treatmentDocumento4 páginasFatal pulmonary leukostasis following AML treatmentkaremiaAún no hay calificaciones
- HEREDITARY RED BLOOD CELL MEMBRANE DISORDERS: SPHEROCYTOSIS, ELLIPTOCYTOSIS, PYROPOIKILOCYTOSIS & STOMATOCYTOSISDocumento29 páginasHEREDITARY RED BLOOD CELL MEMBRANE DISORDERS: SPHEROCYTOSIS, ELLIPTOCYTOSIS, PYROPOIKILOCYTOSIS & STOMATOCYTOSISKristine Mae D. GregorioAún no hay calificaciones
- Diferencias Entre Anemias y TalasemiaDocumento6 páginasDiferencias Entre Anemias y TalasemiaAndreaFonsecaAún no hay calificaciones
- Patholo Le2Documento31 páginasPatholo Le2bitangyarahAún no hay calificaciones
- 1 s2.0 S000649712300366X MainDocumento12 páginas1 s2.0 S000649712300366X MainlaksmitatetanelAún no hay calificaciones
- Hematologic System LectureDocumento15 páginasHematologic System LecturedumonilaAún no hay calificaciones
- Harvard BusinessDocumento6 páginasHarvard BusinessACAún no hay calificaciones
- megaloblastic_anemia__an_updated_review.5Documento4 páginasmegaloblastic_anemia__an_updated_review.5Hafizh MusyafaAún no hay calificaciones
- Blood Chemistry - Dra. Gabaldon (RECT) PDFDocumento7 páginasBlood Chemistry - Dra. Gabaldon (RECT) PDFRachel AsuncionAún no hay calificaciones
- Medical Progress: Review ArticlesDocumento11 páginasMedical Progress: Review ArticlesEllie satrianiAún no hay calificaciones
- A New Way To Prevent Thrombosis - 2007 - BloodDocumento1 páginaA New Way To Prevent Thrombosis - 2007 - BloodTareeqanwar MohammedAún no hay calificaciones
- Platelet A-GranulesDocumento11 páginasPlatelet A-GranulesTahir AliAún no hay calificaciones
- Mahajan 2016Documento10 páginasMahajan 2016Juan RamirezAún no hay calificaciones
- Red Blood Cell Disorders Part 1 and 2Documento7 páginasRed Blood Cell Disorders Part 1 and 2babael.zuher123Aún no hay calificaciones
- Hoban 2016 Blood - Genetic Treatment of A Molecular Disorder Gene Therapy Approaches To Sickle Cell DiseaseDocumento11 páginasHoban 2016 Blood - Genetic Treatment of A Molecular Disorder Gene Therapy Approaches To Sickle Cell DiseaseNicolas HEBERTAún no hay calificaciones
- PLT DisordersDocumento7 páginasPLT DisordersAbby CastilloAún no hay calificaciones
- Mechanisms of Disease: V - B - S H H SDocumento10 páginasMechanisms of Disease: V - B - S H H SMedranoReyesLuisinAún no hay calificaciones
- Hematology Week 4 PDFDocumento9 páginasHematology Week 4 PDFKenj Yolleth IbañezAún no hay calificaciones
- Hematopoiesis: 6 - Blood andDocumento38 páginasHematopoiesis: 6 - Blood andRuth MSAún no hay calificaciones
- Hema 2 - Prelim Topic 3 - Platelet Disorders and Laboratory TestsDocumento8 páginasHema 2 - Prelim Topic 3 - Platelet Disorders and Laboratory TestsLowenstein JenzenAún no hay calificaciones
- Molecular Detection of BCR-ABL Fusion GeneDocumento14 páginasMolecular Detection of BCR-ABL Fusion GeneNimra ZulfiqarAún no hay calificaciones
- Cerebrospinal Fluid - AUBFDocumento9 páginasCerebrospinal Fluid - AUBFMitch IbayAún no hay calificaciones
- Desordenes de Membrana InfDocumento3 páginasDesordenes de Membrana InfwillmedAún no hay calificaciones
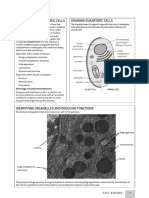
- Structure of Eukaryotic Cells Drawing Eukaryotic CellsDocumento4 páginasStructure of Eukaryotic Cells Drawing Eukaryotic CellsAldo LoyolaAún no hay calificaciones
- Coagulation Falanga 4Documento1 páginaCoagulation Falanga 4s3lki3Aún no hay calificaciones
- Hematology White Cell DisordersDocumento5 páginasHematology White Cell DisordersIS99057Aún no hay calificaciones
- Red Cell Membrane and Abo Blood Group SystemDocumento37 páginasRed Cell Membrane and Abo Blood Group SystemGracia Dela CruzAún no hay calificaciones
- Vivarelli 2019Documento4 páginasVivarelli 2019chloe1411Aún no hay calificaciones
- HEMOGLOBINOPATHY GUIDEDocumento3 páginasHEMOGLOBINOPATHY GUIDEChatie PipitAún no hay calificaciones
- Jacob Et Al 2018 More Than Meets The EyeDocumento4 páginasJacob Et Al 2018 More Than Meets The Eyemmartinezs2507Aún no hay calificaciones
- Pathophysiology of Anemia: A. Gerson Greenburg, MD, PHDDocumento5 páginasPathophysiology of Anemia: A. Gerson Greenburg, MD, PHDAlma AwaliyahAún no hay calificaciones
- Osmosis 44Documento1 páginaOsmosis 44Sarawini KosolsombatAún no hay calificaciones
- The glomerular basement membrane: not gone, just forgottenDocumento4 páginasThe glomerular basement membrane: not gone, just forgottenalma ramirezAún no hay calificaciones
- Midterm Long Test ReviewDocumento2 páginasMidterm Long Test ReviewCarl DevinAún no hay calificaciones
- Feline Nonregenerative Anemia Pathophysiology and ErilologiesDocumento7 páginasFeline Nonregenerative Anemia Pathophysiology and ErilologiesTORRES GUTIERREZ JOYNY RAQUEL catedraticoAún no hay calificaciones
- Cerebrospinal Fluid: Prepared By: Muath M. Abu-Samreh Supervisor: Dr. Samir KhalilDocumento12 páginasCerebrospinal Fluid: Prepared By: Muath M. Abu-Samreh Supervisor: Dr. Samir KhalilMohammad Abu-SamrehAún no hay calificaciones
- Red Cell: Blood GlycophorinsDocumento11 páginasRed Cell: Blood GlycophorinsChu NgọcAún no hay calificaciones
- Cerebrospinal FluidDocumento12 páginasCerebrospinal FluidMuath M. AbusamrehAún no hay calificaciones
- Detecting and Identifying RBC AntibodiesDocumento5 páginasDetecting and Identifying RBC AntibodiesJoy ZapantaAún no hay calificaciones
- Patho HematologyDocumento39 páginasPatho HematologyCastleKGAún no hay calificaciones
- Organelle/ Inclusion Functions LM Features EM Features Associated DiseasesDocumento6 páginasOrganelle/ Inclusion Functions LM Features EM Features Associated DiseasesAine CebedoAún no hay calificaciones
- Imuno Sero Lecture 2Documento16 páginasImuno Sero Lecture 2Marjorie Balangue MacadaegAún no hay calificaciones
- Esplenectomía en EsferocitosisDocumento16 páginasEsplenectomía en EsferocitosisDra_MaradyAún no hay calificaciones
- LEC AUBF Renal-Diseases MIDTERMS 02Documento3 páginasLEC AUBF Renal-Diseases MIDTERMS 02Jashmine May TadinaAún no hay calificaciones
- Anemia Pernicious Anemia Aplastic Anemia Iron Deficiency Anemia Sickle Cell Anemia Hypovolemic (Blood Loss) AnemiaDocumento1 páginaAnemia Pernicious Anemia Aplastic Anemia Iron Deficiency Anemia Sickle Cell Anemia Hypovolemic (Blood Loss) AnemiaJona Mae Ludovice SalonoyAún no hay calificaciones
- Bacterial Genetic SystemDocumento31 páginasBacterial Genetic SystemBatool AshrafAún no hay calificaciones
- Glycoproteins of Cell Surfaces: A Comparative StudyDocumento9 páginasGlycoproteins of Cell Surfaces: A Comparative StudyAdrianacalderonAún no hay calificaciones
- An Outline of Essential Topics in Glomerular Pathophysiology, Diagnosis, and Treatment For Nephrology TraineesDocumento24 páginasAn Outline of Essential Topics in Glomerular Pathophysiology, Diagnosis, and Treatment For Nephrology TraineesKity SarkarAún no hay calificaciones
- Introduction of Natural ImmunityDocumento4 páginasIntroduction of Natural ImmunitySamanthaAún no hay calificaciones
- Isolation of Lymphoid Cells: - Antigen-Antibody Interactions - Antigen-Antibody Based ExperimentsDocumento111 páginasIsolation of Lymphoid Cells: - Antigen-Antibody Interactions - Antigen-Antibody Based ExperimentsSivarama KrishnanAún no hay calificaciones
- Hematology Overview and RBC AnalysisDocumento9 páginasHematology Overview and RBC AnalysisMonica DomingoAún no hay calificaciones
- Labs SVDocumento10 páginasLabs SVLica_Zurriaga_9142Aún no hay calificaciones
- Thrombolytic Therapy in Emergency MedicineDocumento9 páginasThrombolytic Therapy in Emergency MedicinemedeviAún no hay calificaciones
- HHS Public Access: Structure and Function of The Human Skin MicrobiomeDocumento20 páginasHHS Public Access: Structure and Function of The Human Skin MicrobiomeEugeniaAún no hay calificaciones
- Protected Area Expansion Strategy For CapeNature 2010 29 SeptemberDocumento70 páginasProtected Area Expansion Strategy For CapeNature 2010 29 SeptembertableviewAún no hay calificaciones
- Module 2 NCM 114Documento11 páginasModule 2 NCM 114Erven AranasAún no hay calificaciones
- General Biology 2 Budget of WorkDocumento3 páginasGeneral Biology 2 Budget of WorkMaricris BalboaAún no hay calificaciones
- The Science of NeglectDocumento20 páginasThe Science of NeglectdrdrtsaiAún no hay calificaciones
- Crop Sci 1 Lecture Manual OverviewDocumento86 páginasCrop Sci 1 Lecture Manual OverviewAnalYn Litawan Bucasan100% (1)
- Flowering PlantsDocumento43 páginasFlowering Plantskingbanakon100% (1)
- Field Inspection Techniques Ifad-CaspDocumento17 páginasField Inspection Techniques Ifad-CaspNasiru Kura100% (1)
- COT TRF Science G8 - Food ChainDocumento8 páginasCOT TRF Science G8 - Food ChainRAMIR BECOYAún no hay calificaciones
- Biology of Implant OsseointegrationDocumento11 páginasBiology of Implant OsseointegrationAmIrAún no hay calificaciones
- C. S. Sureka, Christina Armpilia - Radiation Biology For Medical Physicists (2017, CRC Press, Taylor & Francis Group) - 148-163Documento16 páginasC. S. Sureka, Christina Armpilia - Radiation Biology For Medical Physicists (2017, CRC Press, Taylor & Francis Group) - 148-163dwi riris indriyaniAún no hay calificaciones
- Lesson 7 - Organ TransplantationDocumento14 páginasLesson 7 - Organ TransplantationJosee100% (1)
- Checkerboard Microdilution AssayDocumento5 páginasCheckerboard Microdilution AssayClaudio Luis Venturini0% (1)
- Development of The Planet EarthDocumento14 páginasDevelopment of The Planet EarthHana CpnplnAún no hay calificaciones
- SOP EcoliDocumento3 páginasSOP EcoliIeqa HaziqahAún no hay calificaciones
- Explaining Butterfly MetamorphosisDocumento13 páginasExplaining Butterfly MetamorphosisCipe drAún no hay calificaciones
- NSABB Final Report Recommendations Evaluation Oversight Proposed Gain of Function Research May 2016Documento109 páginasNSABB Final Report Recommendations Evaluation Oversight Proposed Gain of Function Research May 2016Jim Hoft100% (1)
- Citric Acid Production Stirred TankDocumento9 páginasCitric Acid Production Stirred TankKarliiux MedinaAún no hay calificaciones
- Everyday Sports Injuries The Essential Step-By-Step Guide To Prevention, Diagnosis, and TreatmentDocumento274 páginasEveryday Sports Injuries The Essential Step-By-Step Guide To Prevention, Diagnosis, and TreatmentAnonymous f6goFflg3T100% (10)
- 421 Reproduction Developmental Toxicity Screening TestDocumento10 páginas421 Reproduction Developmental Toxicity Screening TestAshishAún no hay calificaciones
- Summary of Qualitative Tests (Pharmacognosy)Documento8 páginasSummary of Qualitative Tests (Pharmacognosy)kidsaintfineAún no hay calificaciones
- Chapter 5Documento14 páginasChapter 5Caitlin G.Aún no hay calificaciones
- Noscapine - British PharmacopoeiaDocumento3 páginasNoscapine - British PharmacopoeiaSocial Service (V)Aún no hay calificaciones
- Met. Secundarios Cáscara SagradaDocumento9 páginasMet. Secundarios Cáscara SagradaDalíAsesoríasAún no hay calificaciones
- Paul Atriedes (PL 6/181pp) : Bene Gesserit Training (Container, Active 11)Documento2 páginasPaul Atriedes (PL 6/181pp) : Bene Gesserit Training (Container, Active 11)dfgdfAún no hay calificaciones
- Advanced Level Biology 2015 Marking SchemeDocumento29 páginasAdvanced Level Biology 2015 Marking SchemeAngiee FAún no hay calificaciones
- Analisis Kualitatif Dan Kuantitatif Kandungan Kimia Dari Ekstrak Heksan, Aseton, Etanol Dan Air Dari Umbi Bawang Putih (Allium Sativum Linn.)Documento11 páginasAnalisis Kualitatif Dan Kuantitatif Kandungan Kimia Dari Ekstrak Heksan, Aseton, Etanol Dan Air Dari Umbi Bawang Putih (Allium Sativum Linn.)Tari PratiwiAún no hay calificaciones
- Young Children's Biological Predisposition To Learn in Privileged DomainDocumento6 páginasYoung Children's Biological Predisposition To Learn in Privileged DomainVeronica Dadal0% (1)
- Human Dimensions of Late Pleistocene/Holocene Arid Events in Southern South AmericaDocumento13 páginasHuman Dimensions of Late Pleistocene/Holocene Arid Events in Southern South AmericaKaren RochaAún no hay calificaciones
- Is That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeDe EverandIs That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Stuff Matters: Exploring the Marvelous Materials That Shape Our Man-Made WorldDe EverandStuff Matters: Exploring the Marvelous Materials That Shape Our Man-Made WorldCalificación: 4 de 5 estrellas4/5 (289)
- Chemistry for Breakfast: The Amazing Science of Everyday LifeDe EverandChemistry for Breakfast: The Amazing Science of Everyday LifeCalificación: 4.5 de 5 estrellas4.5/5 (14)
- Guidelines for Asset Integrity ManagementDe EverandGuidelines for Asset Integrity ManagementCalificación: 5 de 5 estrellas5/5 (1)
- Monkeys, Myths, and Molecules: Separating Fact from Fiction in the Science of Everyday LifeDe EverandMonkeys, Myths, and Molecules: Separating Fact from Fiction in the Science of Everyday LifeCalificación: 4 de 5 estrellas4/5 (9)
- The Disappearing Spoon: And Other True Tales of Madness, Love, and the History of the World from the Periodic Table of the ElementsDe EverandThe Disappearing Spoon: And Other True Tales of Madness, Love, and the History of the World from the Periodic Table of the ElementsCalificación: 4 de 5 estrellas4/5 (146)
- Science Goes Viral: Captivating Accounts of Science in Everyday LifeDe EverandScience Goes Viral: Captivating Accounts of Science in Everyday LifeCalificación: 5 de 5 estrellas5/5 (1)
- Coating and Drying Defects: Troubleshooting Operating ProblemsDe EverandCoating and Drying Defects: Troubleshooting Operating ProblemsCalificación: 5 de 5 estrellas5/5 (1)
- Organic Chemistry for Schools: Advanced Level and Senior High SchoolDe EverandOrganic Chemistry for Schools: Advanced Level and Senior High SchoolAún no hay calificaciones
- An Introduction to the Periodic Table of Elements : Chemistry Textbook Grade 8 | Children's Chemistry BooksDe EverandAn Introduction to the Periodic Table of Elements : Chemistry Textbook Grade 8 | Children's Chemistry BooksCalificación: 5 de 5 estrellas5/5 (1)
- Guidelines for Defining Process Safety Competency RequirementsDe EverandGuidelines for Defining Process Safety Competency RequirementsCalificación: 3 de 5 estrellas3/5 (1)
- Napoleon's Buttons: 17 Molecules That Changed HistoryDe EverandNapoleon's Buttons: 17 Molecules That Changed HistoryCalificación: 4 de 5 estrellas4/5 (25)
- The Regenerative Grower's Guide to Garden Amendments: Using Locally Sourced Materials to Make Mineral and Biological Extracts and FermentsDe EverandThe Regenerative Grower's Guide to Garden Amendments: Using Locally Sourced Materials to Make Mineral and Biological Extracts and FermentsCalificación: 5 de 5 estrellas5/5 (3)
- Essential Oil Chemistry Formulating Essential Oil Blends that Heal - Aldehyde - Ketone - Lactone: Healing with Essential OilDe EverandEssential Oil Chemistry Formulating Essential Oil Blends that Heal - Aldehyde - Ketone - Lactone: Healing with Essential OilCalificación: 5 de 5 estrellas5/5 (1)
- It's Elemental: The Hidden Chemistry in EverythingDe EverandIt's Elemental: The Hidden Chemistry in EverythingCalificación: 4 de 5 estrellas4/5 (10)
- Meltdown: Nuclear disaster and the human cost of going criticalDe EverandMeltdown: Nuclear disaster and the human cost of going criticalCalificación: 5 de 5 estrellas5/5 (5)
- Chemical Elements Pocket Guide: Detailed Summary of the Periodic TableDe EverandChemical Elements Pocket Guide: Detailed Summary of the Periodic TableAún no hay calificaciones
- Chemistry: a QuickStudy Laminated Reference GuideDe EverandChemistry: a QuickStudy Laminated Reference GuideCalificación: 5 de 5 estrellas5/5 (1)
- The Periodic Table: A Very Short IntroductionDe EverandThe Periodic Table: A Very Short IntroductionCalificación: 4.5 de 5 estrellas4.5/5 (3)
- Chemistry for Breakfast: The Amazing Science of Everyday LifeDe EverandChemistry for Breakfast: The Amazing Science of Everyday LifeCalificación: 4.5 de 5 estrellas4.5/5 (90)
- The Elements We Live By: How Iron Helps Us Breathe, Potassium Lets Us See, and Other Surprising Superpowers of the Periodic TableDe EverandThe Elements We Live By: How Iron Helps Us Breathe, Potassium Lets Us See, and Other Surprising Superpowers of the Periodic TableCalificación: 3.5 de 5 estrellas3.5/5 (22)
- Introduction to Strategies for Organic SynthesisDe EverandIntroduction to Strategies for Organic SynthesisAún no hay calificaciones
- Guidelines for Integrating Process Safety into Engineering ProjectsDe EverandGuidelines for Integrating Process Safety into Engineering ProjectsAún no hay calificaciones
- Monkeys, Myths, and Molecules: Separating Fact from Fiction, and the Science of Everyday LifeDe EverandMonkeys, Myths, and Molecules: Separating Fact from Fiction, and the Science of Everyday LifeCalificación: 4 de 5 estrellas4/5 (1)