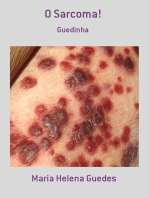
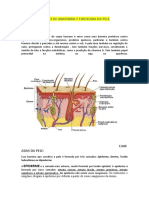
También podría gustarte
- O Papel Do Farmacêutico Na Cicatrização De FeridasDe EverandO Papel Do Farmacêutico Na Cicatrização De FeridasAún no hay calificaciones
- Eletroterapia - LASER TerapêuticoDocumento81 páginasEletroterapia - LASER TerapêuticoJosé Carlos100% (1)
- Protocolo Cief HCM PDFDocumento45 páginasProtocolo Cief HCM PDFDejair RodolfiAún no hay calificaciones
- A regulação da cosmiatria no Brasil: o conflito regulatório entre os conselhos profissionais de saúdeDe EverandA regulação da cosmiatria no Brasil: o conflito regulatório entre os conselhos profissionais de saúdeAún no hay calificaciones
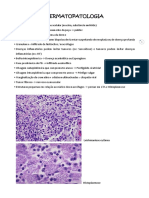
- DERMATOPATOLOGIADocumento72 páginasDERMATOPATOLOGIAMiguel Ceccarelli CalleAún no hay calificaciones
- CarboxiDocumento18 páginasCarboxiRose FigueiredoAún no hay calificaciones
- Receptores Sensoriais TransparenciaDocumento6 páginasReceptores Sensoriais TransparenciaDouglasVizzuAún no hay calificaciones
- Semiologia Da PeleDocumento4 páginasSemiologia Da PelePhoenix ImmortalAún no hay calificaciones
- Cosmetologia Corporal AplicadaDocumento11 páginasCosmetologia Corporal AplicadaLeonardo BarrosAún no hay calificaciones
- Proteinas e EnzimasDocumento19 páginasProteinas e EnzimasApolo100% (1)
- EscabioseDocumento7 páginasEscabiosepepeladazoAún no hay calificaciones
- Apostila EnvelhecimentoDocumento19 páginasApostila Envelhecimentocris100% (1)
- DERMEDocumento44 páginasDERMEMariana Nóbrega La MarcaAún no hay calificaciones
- Tarefa Avaliativa - Bruno e Luís GustavoDocumento14 páginasTarefa Avaliativa - Bruno e Luís GustavoBruno Marques100% (1)
- Pele e Anexos - Histologia InterativaDocumento11 páginasPele e Anexos - Histologia Interativarenata de souza100% (1)
- Infeccoes Fungicas Final OkDocumento6 páginasInfeccoes Fungicas Final OkHyo SilvaAún no hay calificaciones
- Livro 4 Ciencia e Tecnologia em SaudeDocumento168 páginasLivro 4 Ciencia e Tecnologia em SaudeGeovani Paz Rodrigues MirandaAún no hay calificaciones
- Entendendo A Alopecia AndrogenéticaDocumento10 páginasEntendendo A Alopecia AndrogenéticaAlexia Vieira de SouzaAún no hay calificaciones
- PRPDocumento4 páginasPRPNR_BinhaAún no hay calificaciones
- Estrategias Terapeuticas No Manejo Do FegDocumento4 páginasEstrategias Terapeuticas No Manejo Do FegJuliana SantanaAún no hay calificaciones
- E-Book o Que Fazer Com As HipercromiasDocumento39 páginasE-Book o Que Fazer Com As HipercromiasSonia Mendonça CalixtoAún no hay calificaciones
- Cosmeticos GestantesDocumento10 páginasCosmeticos GestantesTatianaAún no hay calificaciones
- Foliculite - ProntoDocumento12 páginasFoliculite - ProntoErica TadianeAún no hay calificaciones
- Revisão Anátomo Fisiológica Do Sistema Tegumentar PDFDocumento81 páginasRevisão Anátomo Fisiológica Do Sistema Tegumentar PDFJuliana Pontes100% (1)
- Manual de DermatopatologiaDocumento36 páginasManual de DermatopatologiaMarcelo Lima100% (2)
- A Pele (Aula Completa)Documento123 páginasA Pele (Aula Completa)Anna PinheiroAún no hay calificaciones
- Sistema Tegumentar LesõesDocumento12 páginasSistema Tegumentar LesõeslucianolimaglobalAún no hay calificaciones
- Redução Do Edema em Membros Inferiores Através Da DrenagemDocumento49 páginasRedução Do Edema em Membros Inferiores Através Da Drenagemmarciokh100% (5)
- Descomplicando o Sistema Tegumentar PDFDocumento51 páginasDescomplicando o Sistema Tegumentar PDFDaianeRochaEsteticistaAún no hay calificaciones
- Anatomia e Fisiologia Da Pele - Sem FotosDocumento53 páginasAnatomia e Fisiologia Da Pele - Sem Fotosamandinhabm_18Aún no hay calificaciones
- MelasmaDocumento13 páginasMelasmaRose FigueiredoAún no hay calificaciones
- Crescimento e Renovação CelularDocumento7 páginasCrescimento e Renovação CelularJoana TeixeiraAún no hay calificaciones
- Caracterização Do Fibro Edema Gelóide e PDFDocumento59 páginasCaracterização Do Fibro Edema Gelóide e PDFJanneAún no hay calificaciones
- Curso de DepilaçãoDocumento13 páginasCurso de DepilaçãoJessica Pinheiro EsteticaAún no hay calificaciones
- Resumo DermatoDocumento86 páginasResumo DermatoSuzana CarvalhoAún no hay calificaciones
- Apostila SoroterapiaDocumento18 páginasApostila Soroterapiapsicanalistaanaesteves100% (1)
- Celulite - TrabalhoDocumento10 páginasCelulite - TrabalhoLarissa RochaAún no hay calificaciones
- Estetica e Imagem Pessoal 2Documento40 páginasEstetica e Imagem Pessoal 2Marcelo BrunoAún no hay calificaciones
- 02 Apostila Funcoes - Da - PeleDocumento16 páginas02 Apostila Funcoes - Da - PeleAline100% (1)
- Pele e Anexos PDFDocumento46 páginasPele e Anexos PDF01Jessica02100% (1)
- Patologia Do Sistema TegumentarDocumento8 páginasPatologia Do Sistema TegumentarBeatrizAún no hay calificaciones
- Óleo de Melaleuca No Tratamento Da AcneDocumento10 páginasÓleo de Melaleuca No Tratamento Da AcneIrys LopesAún no hay calificaciones
- ARTIGO 1 - O Uso Associado Do Ácido Kójico e Ácido Glicólico Como Alternativa À Hidroquinona No Tratamento de MelasmaDocumento15 páginasARTIGO 1 - O Uso Associado Do Ácido Kójico e Ácido Glicólico Como Alternativa À Hidroquinona No Tratamento de MelasmaTertaliana PiresAún no hay calificaciones
- Protocolo UpGluteos 195x69mmDocumento1 páginaProtocolo UpGluteos 195x69mmThatiana RibeiroAún no hay calificaciones
- Ebook OzonioOxDocumento32 páginasEbook OzonioOxcarolinaAún no hay calificaciones
- Sistema Tegumentar - Alteraçoes AnatomicasDocumento50 páginasSistema Tegumentar - Alteraçoes AnatomicasZoeAún no hay calificaciones
- Curso Limpeza de Pele OkDocumento17 páginasCurso Limpeza de Pele OkErica Tadiane100% (1)
- Associação Entre Fototerapia e Terapia Fotodinâmica No Tratamento de Ferida Cutânea em CãoDocumento7 páginasAssociação Entre Fototerapia e Terapia Fotodinâmica No Tratamento de Ferida Cutânea em CãoFairy LudaAún no hay calificaciones
- Diagnostico de Hiperadrenocorticismo Canino e Tratamento Com TrilostanoDocumento148 páginasDiagnostico de Hiperadrenocorticismo Canino e Tratamento Com TrilostanoGlauce L Trevisan100% (1)
- Apostila Curso Online Limpeza de PeleDocumento72 páginasApostila Curso Online Limpeza de PeleDeyvison MendesAún no hay calificaciones
- Dermatologia - 4A MIII A13 - AlopeciasDocumento12 páginasDermatologia - 4A MIII A13 - AlopeciasFELIPE EMANUEL BASNIAK DA SILVA (A)Aún no hay calificaciones
- Atm - Articulação TemporomandibularDocumento51 páginasAtm - Articulação TemporomandibularRodrigo OliveiraAún no hay calificaciones
- Tabela de DermatitesDocumento7 páginasTabela de DermatitesAgatha RedAún no hay calificaciones
- FotoproteçãoDocumento7 páginasFotoproteçãoRaquel Ávila Nozari100% (1)
- #ParasitologiaDocumento32 páginas#ParasitologiaKarinaAún no hay calificaciones
- Manual de IontoforeseDocumento72 páginasManual de IontoforeseHildemar ShirataAún no hay calificaciones
- E-Book Lobuloplastia Não CirúrgicaDocumento31 páginasE-Book Lobuloplastia Não CirúrgicaGiovana PeronAún no hay calificaciones
- Micropuntura - Rafael FerreiraDocumento1 páginaMicropuntura - Rafael FerreiraClaudio InoveAún no hay calificaciones
- CitocinasDocumento11 páginasCitocinasSuzana PoloncaAún no hay calificaciones
- Priscilaferrari Final 150628021110 Lva1 App6891Documento45 páginasPriscilaferrari Final 150628021110 Lva1 App6891Suzana Polonca50% (2)
- v9 Conceitos Atuais No Uso Do Acido Poli L Lactico para Rejuvenescimento Facial Revisao e Aspectos PraticosDocumento12 páginasv9 Conceitos Atuais No Uso Do Acido Poli L Lactico para Rejuvenescimento Facial Revisao e Aspectos PraticosSuzana PoloncaAún no hay calificaciones
- Mariana Robatto Dantas LealDocumento55 páginasMariana Robatto Dantas LealSuzana PoloncaAún no hay calificaciones
- 4 Radiofrequnciamultifrequencialemultipolarnostratamentoscorporaisefaciais 140924154343 Phpapp02Documento51 páginas4 Radiofrequnciamultifrequencialemultipolarnostratamentoscorporaisefaciais 140924154343 Phpapp02Suzana Polonca100% (3)
- 201411584948148ppc Tecnico em Estetica Integrado Proeja - SanDocumento69 páginas201411584948148ppc Tecnico em Estetica Integrado Proeja - SanSuzana PoloncaAún no hay calificaciones
- Conheça o Processo Básico para Se Formar Uma Associação e Quais As EtapasDocumento32 páginasConheça o Processo Básico para Se Formar Uma Associação e Quais As EtapasSuzana PoloncaAún no hay calificaciones
- Guia Oncologia BiooncoDocumento22 páginasGuia Oncologia BiooncoSuzana PoloncaAún no hay calificaciones
- Perimetria e Dobras Cutaneas e ProtocolosDocumento50 páginasPerimetria e Dobras Cutaneas e ProtocolosSuzana Polonca100% (1)
- Cosmeceuticos - Bom MaterialDocumento38 páginasCosmeceuticos - Bom MaterialSuzana Polonca100% (1)
- Drug Delivery Fabio BorgesDocumento6 páginasDrug Delivery Fabio BorgesSuzana PoloncaAún no hay calificaciones
- v5 Comparacao Entre Acido Tioglicolico 2 5 Hidroquinona 2 Haloxyl 2 e Peeling de Acido Glicolico 10 No Tratamento Da Hiperpigmentacao PeriorbitalDocumento6 páginasv5 Comparacao Entre Acido Tioglicolico 2 5 Hidroquinona 2 Haloxyl 2 e Peeling de Acido Glicolico 10 No Tratamento Da Hiperpigmentacao PeriorbitalSuzana PoloncaAún no hay calificaciones
- Projeto LiteraturaDocumento6 páginasProjeto LiteraturaFernando RodriguesAún no hay calificaciones
- Flygt 3069Documento3 páginasFlygt 3069Diego Bertelli CuzzuolAún no hay calificaciones
- A Invenção Da ImprensaDocumento3 páginasA Invenção Da ImprensaJoão Felipe Alves de OliveiraAún no hay calificaciones
- Movimento Circular UniformeDocumento15 páginasMovimento Circular UniformeMJTours LdaAún no hay calificaciones
- Atividade Complementar - 6º Ano - 1º BimestreDocumento19 páginasAtividade Complementar - 6º Ano - 1º BimestreFernanda De Souza Alves De JesusAún no hay calificaciones
- Psicopatologia - Mapas Mentais-1626464781-8Documento1 páginaPsicopatologia - Mapas Mentais-1626464781-8Laura GonçalvesAún no hay calificaciones
- Cox (2009,75-90) Estudos Linguísticos MTDocumento16 páginasCox (2009,75-90) Estudos Linguísticos MTJessyca PachecoAún no hay calificaciones
- Caderno de Provas - 2 EtapaDocumento30 páginasCaderno de Provas - 2 EtapaIsadora CoutinhoAún no hay calificaciones
- Supersticao BrasileiraDocumento3 páginasSupersticao BrasileiraJosé Del Carmen EliseoAún no hay calificaciones
- Atividades Não Presenciais 3 Ano - 1 A 5 de FevDocumento7 páginasAtividades Não Presenciais 3 Ano - 1 A 5 de FevFabiolla PiresAún no hay calificaciones
- Manual Do Mel 2 Edicao Web PDFDocumento216 páginasManual Do Mel 2 Edicao Web PDFGina Lecomte0% (1)
- 2.1 - Métodos HistológicosDocumento64 páginas2.1 - Métodos HistológicosHeitor BoraliAún no hay calificaciones
- Tomo VII PDFDocumento857 páginasTomo VII PDFgsterrinha50% (2)
- Questões David HUmeDocumento11 páginasQuestões David HUmeadxaddaAún no hay calificaciones
- Modelo Receituário Especial para ImpressãoDocumento1 páginaModelo Receituário Especial para ImpressãoLetícia Melo100% (1)
- Biologia ForenseDocumento9 páginasBiologia ForenseErillene FurtadoAún no hay calificaciones
- Kendra Little - A Armadilha Do Namorado Bilionário - Família Kavanagh 01Documento109 páginasKendra Little - A Armadilha Do Namorado Bilionário - Família Kavanagh 01Rubi Ariadne100% (2)
- Como Fazer Um Arranjo para Instrumentos de SoproDocumento6 páginasComo Fazer Um Arranjo para Instrumentos de SoproPatrick Ferreira AraujoAún no hay calificaciones
- Atividade de Nota PromissóriaDocumento4 páginasAtividade de Nota PromissóriaMARLON Bruno BarbosaAún no hay calificaciones
- Nit Dicla 16 - 01Documento10 páginasNit Dicla 16 - 01Marcus HugenneyerAún no hay calificaciones
- Planilha de Equalização - Rev02Documento1 páginaPlanilha de Equalização - Rev02Devide LimaAún no hay calificaciones
- Protocolos e Métodos de Análise em Laboratórios de Biotec Agroalimentar e Saúde Humana PDFDocumento330 páginasProtocolos e Métodos de Análise em Laboratórios de Biotec Agroalimentar e Saúde Humana PDFLarissa MAún no hay calificaciones
- Itaucard - 4078 - Fatura - 2024-04Documento4 páginasItaucard - 4078 - Fatura - 2024-04EL Gráfica SolutionAún no hay calificaciones
- Atelier de Projeto de Arquitetura I - Simulado IDocumento4 páginasAtelier de Projeto de Arquitetura I - Simulado ImarcleimvsAún no hay calificaciones
- Relatividade A Passagem Do Enfoque Galileano para A Visão de EinsteinDocumento68 páginasRelatividade A Passagem Do Enfoque Galileano para A Visão de EinsteinPedro MateusAún no hay calificaciones
- A Erótica e o FemininoDocumento186 páginasA Erótica e o FemininoMatheus HenriqueAún no hay calificaciones
- Aborto SeminárioDocumento8 páginasAborto SeminárioMikeAún no hay calificaciones
- Medidas TendenciaCentral ExcelDocumento2 páginasMedidas TendenciaCentral ExcelAndré XavierAún no hay calificaciones
- N 0111Documento41 páginasN 0111Carlosjr83Aún no hay calificaciones
- Caso Clínico Po SupraespinhosoDocumento27 páginasCaso Clínico Po SupraespinhosoCristina CardosoAún no hay calificaciones