También podría gustarte
- Especialidad de FloresDocumento21 páginasEspecialidad de FloresJose Richard Tenazoa VillalobosAún no hay calificaciones
- Fundamentos y técnicas básicas en biología molecularDe EverandFundamentos y técnicas básicas en biología molecularAún no hay calificaciones
- DIAGNÓSTICO DE VERTICALIDAD Y REDONDEZ EN TANQUES DE COMBUSTIBLE POR GEODESIADocumento8 páginasDIAGNÓSTICO DE VERTICALIDAD Y REDONDEZ EN TANQUES DE COMBUSTIBLE POR GEODESIACristian Abanto LeónAún no hay calificaciones
- Los inicios de la vida: La evolución en la Tierra precámbricaDe EverandLos inicios de la vida: La evolución en la Tierra precámbricaAún no hay calificaciones
- Están Vivos Ahora... Por V.P. WierwilleDocumento53 páginasEstán Vivos Ahora... Por V.P. WierwilleOscar Avila100% (6)
- Guía del abogado para el uso de pruebas forenses de ADNDe EverandGuía del abogado para el uso de pruebas forenses de ADNAún no hay calificaciones
- Fuego griego, flechas envenenadas y escorpiones: Guerra química y bacteriológica en la AntigüedadDe EverandFuego griego, flechas envenenadas y escorpiones: Guerra química y bacteriológica en la AntigüedadAún no hay calificaciones
- Derecho A La VidaDocumento10 páginasDerecho A La VidaViviana SánchezAún no hay calificaciones
- Muri, Mura, Muda y Los 7 DesperdiciosDocumento9 páginasMuri, Mura, Muda y Los 7 DesperdiciosVictor Hernandez ArchundiaAún no hay calificaciones
- Resumen TranscripcionDocumento6 páginasResumen Transcripcionpamela valenzuelaAún no hay calificaciones
- Resumen TranscripcionDocumento6 páginasResumen Transcripcionpamela valenzuelaAún no hay calificaciones
- Guia de Actividades Hay Ahi AyDocumento2 páginasGuia de Actividades Hay Ahi AyVictoria AmaroAún no hay calificaciones
- Danny El PerroDocumento5 páginasDanny El PerroValentina Muñoz Oyarzún33% (3)
- Modelo de elegibilidad de préstamos hipotecarios con menos deDocumento3 páginasModelo de elegibilidad de préstamos hipotecarios con menos deFlavio SalazarAún no hay calificaciones
- Nietzsche (1886) 14 Significado de La Locura en La Historia de La Moralidad (Aurora) PDFDocumento3 páginasNietzsche (1886) 14 Significado de La Locura en La Historia de La Moralidad (Aurora) PDFLucas Maya CorreaAún no hay calificaciones
- Cultura de MasasDocumento17 páginasCultura de MasasAndrea FelixAún no hay calificaciones
- E. P. Thompson-De La Historia Social A La IrenologíaDocumento27 páginasE. P. Thompson-De La Historia Social A La IrenologíaAnderson PaulAún no hay calificaciones
- La Base Molecular de La Herencia.: Tema 16Documento42 páginasLa Base Molecular de La Herencia.: Tema 16Alejandro Aguayo TalaveraAún no hay calificaciones
- Descubrimiento del ADN como material genéticoDocumento5 páginasDescubrimiento del ADN como material genéticoLuzmarie CastilloAún no hay calificaciones
- à - CIDOS NUCLEICOS y Sã - Ntesis de Proteã - NasDocumento10 páginasà - CIDOS NUCLEICOS y Sã - Ntesis de Proteã - NasMilagros Sofía ValenzuelaAún no hay calificaciones
- Guia Sobre El Adn - Iraima AquinoDocumento4 páginasGuia Sobre El Adn - Iraima AquinoRitaAún no hay calificaciones
- Primera Parte ProyectoDocumento19 páginasPrimera Parte ProyectolagranjasanitariaAún no hay calificaciones
- Ácidos NucleicosDocumento17 páginasÁcidos Nucleicosailyn heidy yanc baylonAún no hay calificaciones
- Adn Erika UpavDocumento42 páginasAdn Erika UpavDamian LopezAún no hay calificaciones
- GUÍA No. 4 GENÉTICA MOLECULAR EL ADNDocumento14 páginasGUÍA No. 4 GENÉTICA MOLECULAR EL ADNnadir steven jimenez rodriguezAún no hay calificaciones
- 2do Bgu Texto BiologiaDocumento1 página2do Bgu Texto BiologiaBriana TorresAún no hay calificaciones
- SEMANA 4 CUARTO UsadaDocumento6 páginasSEMANA 4 CUARTO UsadaLicedAún no hay calificaciones
- 2-1 Experimentos Clásicos, El ADN Como El Material GenéticoDocumento6 páginas2-1 Experimentos Clásicos, El ADN Como El Material GenéticoJULIETH VALENTINA CARVAJAL MACIASAún no hay calificaciones
- Descubriendo El Material HereditarioDocumento3 páginasDescubriendo El Material HereditarioJavier Ignacio Acevedo MorenoAún no hay calificaciones
- El ADN Como Material GenéticoDocumento7 páginasEl ADN Como Material GenéticoEstelaZetinaAún no hay calificaciones
- Experimentos que demostraron que el ADN es el material genéticoDocumento4 páginasExperimentos que demostraron que el ADN es el material genético2kr7kpzcbjAún no hay calificaciones
- Adn-Pdf GeneticaDocumento22 páginasAdn-Pdf GeneticataraAún no hay calificaciones
- Descubrimiento Del Material Genetico EspañolDocumento4 páginasDescubrimiento Del Material Genetico EspañolLya Badillo AlvarezAún no hay calificaciones
- Las Primeras Evidencias Del ADN Como Material GenéticoDocumento2 páginasLas Primeras Evidencias Del ADN Como Material GenéticooscarAún no hay calificaciones
- 4º - Guía Estudio de Información Génica y ProteínasDocumento18 páginas4º - Guía Estudio de Información Génica y Proteínascsavoy_imlpAún no hay calificaciones
- Descubrimiento Del AdnDocumento7 páginasDescubrimiento Del AdnEddy Joel Ramirez GarciaAún no hay calificaciones
- El Camino A La Doble Hélice - CurtisDocumento3 páginasEl Camino A La Doble Hélice - CurtisOsoPandoAún no hay calificaciones
- Experimentos Clásicos - El ADN Como El Material Genético (Artículo) - Khan AcademyDocumento11 páginasExperimentos Clásicos - El ADN Como El Material Genético (Artículo) - Khan AcademyNicolás Reyes QuinterosAún no hay calificaciones
- Charla de BiologíaDocumento4 páginasCharla de BiologíaDaybellis CisnerosAún no hay calificaciones
- RESEÑA HISTÓRICA DEL ADNDocumento2 páginasRESEÑA HISTÓRICA DEL ADNEliana Itati BenitezAún no hay calificaciones
- La Química de La HerenciaDocumento20 páginasLa Química de La HerenciaKatty Jazmin Bohorquez Zuñay100% (1)
- RESUMENDocumento11 páginasRESUMENTifanny Cuellar RamosAún no hay calificaciones
- Cómo se almacena el material genético en el ADNDocumento17 páginasCómo se almacena el material genético en el ADNGabriel RubioAún no hay calificaciones
- ADN. Docx GabyDocumento34 páginasADN. Docx Gabyanon_715473263Aún no hay calificaciones
- Experimentos ClásicosDocumento5 páginasExperimentos ClásicosClaudia Romina Castañeda BulnesAún no hay calificaciones
- Ilovepdf Merged PDFDocumento27 páginasIlovepdf Merged PDFcamila ignaciaAún no hay calificaciones
- Adn Biologia ComúnDocumento11 páginasAdn Biologia Comúnreload8Aún no hay calificaciones
- Replicación ADN experimento GriffithDocumento14 páginasReplicación ADN experimento GriffithmanuelenecpAún no hay calificaciones
- Guia 1 de Biologia 4m Ejercicios para La Introducción A Biología MolecularDocumento10 páginasGuia 1 de Biologia 4m Ejercicios para La Introducción A Biología MolecularCarol Orielly Savoy VallejosAún no hay calificaciones
- Trabajo FinDocumento18 páginasTrabajo FinStefanyVilcapomaMendezAún no hay calificaciones
- Aportaciones ADNDocumento4 páginasAportaciones ADNnataliaAún no hay calificaciones
- Experimentos clásicos que demostraron el ADN como material genéticoDocumento4 páginasExperimentos clásicos que demostraron el ADN como material genéticoClaudia Romina Castañeda BulnesAún no hay calificaciones
- Material Teórico-Tema 9-ADN Como Responsable de La HerenciaDocumento46 páginasMaterial Teórico-Tema 9-ADN Como Responsable de La HerenciaMarcos SkoroplasAún no hay calificaciones
- Historia Del ADNDocumento14 páginasHistoria Del ADNEduardo José Hernández GuevaraAún no hay calificaciones
- Trabajo de Investigacion Bibliografico Sobre El ADNDocumento8 páginasTrabajo de Investigacion Bibliografico Sobre El ADNLuisAún no hay calificaciones
- Tema 2-Biología Molecular PDFDocumento64 páginasTema 2-Biología Molecular PDFRicardoChuliaAún no hay calificaciones
- Maclyn Mccarty Con Francis Crick Y James D. Watson: Vivo) Como en Tubos de Ensayo (In Vitro)Documento2 páginasMaclyn Mccarty Con Francis Crick Y James D. Watson: Vivo) Como en Tubos de Ensayo (In Vitro)edgleydis urbinaAún no hay calificaciones
- ADNDocumento2 páginasADNvincen.nunezAún no hay calificaciones
- El Nacimiento de La Biología MolecularDocumento1 páginaEl Nacimiento de La Biología MolecularMásvale FeriaAún no hay calificaciones
- I Introducción ExperimentosDocumento9 páginasI Introducción ExperimentossergioaantonioAún no hay calificaciones
- Genética Química Clínica TlaxtlaDocumento6 páginasGenética Química Clínica TlaxtlaJosé María SerranoAún no hay calificaciones
- Bases Quimicas HerenciaDocumento7 páginasBases Quimicas Herenciajuancho123934650% (4)
- Bases Quimicas de La HerenciaDocumento15 páginasBases Quimicas de La HerenciaRoswil LeonAún no hay calificaciones
- Avances Científicos en BiologíaDocumento5 páginasAvances Científicos en BiologíaYuriKatherineUribeJimenezAún no hay calificaciones
- Resumen Encontramos El Secreto de La VidaDocumento3 páginasResumen Encontramos El Secreto de La VidaVanessa Díaz0% (1)
- Aportes clave a la biologíaDocumento3 páginasAportes clave a la biologíaLach Fuquencio Chepe PiscoyaAún no hay calificaciones
- Biologia S3 Primera Expo. de CICLO-1Documento24 páginasBiologia S3 Primera Expo. de CICLO-1Pedro Alberto Silva EgusquizaAún no hay calificaciones
- 1 - Historia de La Biología MolecularDocumento7 páginas1 - Historia de La Biología MolecularHUMBERTO ALEJANDRO GARCIA SALAZARAún no hay calificaciones
- ADN Como Material GeneticoDocumento52 páginasADN Como Material GeneticosanquimicoAún no hay calificaciones
- 1869 Friedrich NietzscheDocumento4 páginas1869 Friedrich NietzscheJeffersonDanielArgüelloObandoAún no hay calificaciones
- Genética I (T1 - T13)Documento149 páginasGenética I (T1 - T13)Rocío Prieto VázquezAún no hay calificaciones
- Bmyg MCDocumento135 páginasBmyg MCnicole salazarAún no hay calificaciones
- MembDocumento4 páginasMembpamela valenzuelaAún no hay calificaciones
- Sistema respiratorio: estructuras y funciones claveDocumento9 páginasSistema respiratorio: estructuras y funciones clavepamela valenzuelaAún no hay calificaciones
- Clasificacion TaxonomicaDocumento4 páginasClasificacion Taxonomicapamela valenzuelaAún no hay calificaciones
- Septimo PrimeraDocumento7 páginasSeptimo Primerapamela valenzuelaAún no hay calificaciones
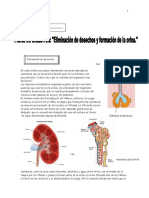
- Eliminación de desechos metabólicos a través de los riñones y la pielDocumento3 páginasEliminación de desechos metabólicos a través de los riñones y la pielpamela valenzuelaAún no hay calificaciones
- EcologiaDocumento8 páginasEcologiapamela valenzuelaAún no hay calificaciones
- Repaso SeptimoDocumento8 páginasRepaso Septimopamela valenzuelaAún no hay calificaciones
- Séptimo ActividadDocumento1 páginaSéptimo Actividadpamela valenzuelaAún no hay calificaciones
- Conociendo Mis EmocionesDocumento7 páginasConociendo Mis EmocionesPastor Agustin Cervantes75% (4)
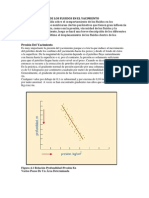
- Comportamiento de Los Fluidos en El YacimientoDocumento15 páginasComportamiento de Los Fluidos en El YacimientoGuerra YilAún no hay calificaciones
- MergedDocumento69 páginasMergedAlan Varela GarcíaAún no hay calificaciones
- Objetivo 3 - Ejercicios Funcion AfinDocumento10 páginasObjetivo 3 - Ejercicios Funcion AfinOmar OrteganoAún no hay calificaciones
- Generar gráfica de 500 números aleatorios LabViewDocumento3 páginasGenerar gráfica de 500 números aleatorios LabViewSergio RamosAún no hay calificaciones
- Carpe DiemDocumento13 páginasCarpe DiemGustavoCruzVegaAún no hay calificaciones
- 2.2 Caso Consolidación Directa Con Ventas AscendentesDocumento20 páginas2.2 Caso Consolidación Directa Con Ventas AscendentesFranks RamirezAún no hay calificaciones
- Glosario y MYPEDocumento2 páginasGlosario y MYPERenzo Carrasco Robles100% (1)
- Ac - ReumatologíaDocumento18 páginasAc - ReumatologíaducAún no hay calificaciones
- Semana N°19 (El Boom Hispanoamericano)Documento3 páginasSemana N°19 (El Boom Hispanoamericano)Carlos M.0% (1)
- Sistemas de Encofrado - Entibación - ES W 02 1211Documento24 páginasSistemas de Encofrado - Entibación - ES W 02 1211Samantha PowellAún no hay calificaciones
- Dependencia y adicción a las benzodiazepinasDocumento9 páginasDependencia y adicción a las benzodiazepinasHelix Ivan Barajas CalderonAún no hay calificaciones
- Manual de Operaciones Del FPSDocumento70 páginasManual de Operaciones Del FPSGojim IngenierosAún no hay calificaciones
- Aderezas Mesa Delante de Mí en Presencia de Mis AngustiadoresDocumento3 páginasAderezas Mesa Delante de Mí en Presencia de Mis AngustiadoresMisión Llano GrandeAún no hay calificaciones
- Espacios-Ana Kely Andrade Q.Documento15 páginasEspacios-Ana Kely Andrade Q.Kely AndradeAún no hay calificaciones
- 00 U2000 R16C50 Funciones BásicasDocumento82 páginas00 U2000 R16C50 Funciones BásicasAlvaro CueAún no hay calificaciones
- Infografía Informativa Salud Sexual Ilustrado Vintage Rosa MentaDocumento1 páginaInfografía Informativa Salud Sexual Ilustrado Vintage Rosa Mentafiorella sirlopuAún no hay calificaciones
- Raz. Verbal 3ero Sec - T1 PDFDocumento4 páginasRaz. Verbal 3ero Sec - T1 PDFFranchesco mendozaAún no hay calificaciones
- Actitud Del ConsumidorDocumento3 páginasActitud Del ConsumidorJonah GalarzaAún no hay calificaciones