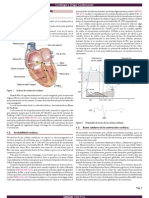
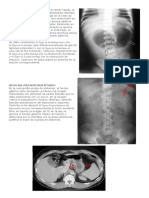
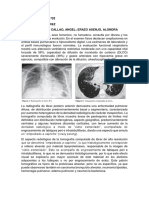
También podría gustarte
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017De EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Aún no hay calificaciones
- Historia Clínica Formato USMPDocumento9 páginasHistoria Clínica Formato USMPMartin Eduardo Marchena TiradoAún no hay calificaciones
- Indice General de Preguntas OncologiaDocumento781 páginasIndice General de Preguntas OncologiaAngel Stevens Garcia Callao100% (2)
- Seminario 4 FisiologiaDocumento17 páginasSeminario 4 FisiologiaAlondra Erazo AsenjoAún no hay calificaciones
- Seminario 4 MemoriaDocumento35 páginasSeminario 4 MemoriaBeatriz VillarrealAún no hay calificaciones
- Potenciación de Largo PlazoDocumento4 páginasPotenciación de Largo PlazoYamila BarberioAún no hay calificaciones
- LA MEMORIA: PROCESOS COGNITIVOS Y NEURONALESDocumento1 páginaLA MEMORIA: PROCESOS COGNITIVOS Y NEURONALESDayanna PalominoAún no hay calificaciones
- Síndromes de Funciones Mentales SuperioresDocumento25 páginasSíndromes de Funciones Mentales SuperioresAVILA ZEPEDA ARMANDOAún no hay calificaciones
- Ciencias Básicas NeurologíaDocumento25 páginasCiencias Básicas NeurologíaBianca DiazAún no hay calificaciones
- Biologia SEMANA 12Documento3 páginasBiologia SEMANA 12john salchichon100% (2)
- Introducción A La Farmacología Del Sistema Nervioso AutónomoDocumento35 páginasIntroducción A La Farmacología Del Sistema Nervioso AutónomoKassandra AranaAún no hay calificaciones
- Liceo Carmela Carvajal de PratDocumento8 páginasLiceo Carmela Carvajal de PratSilvana Selene Egas EgasAún no hay calificaciones
- Tema 6 FisiologicaDocumento7 páginasTema 6 FisiologicaACRAún no hay calificaciones
- Erik Jerson Flores QuispeDocumento13 páginasErik Jerson Flores QuispeErik J. Roshan FloresAún no hay calificaciones
- Seminario 4 - MemoriaDocumento16 páginasSeminario 4 - MemoriaLourdesParedesMedinaAún no hay calificaciones
- Neuro SinapsisDocumento5 páginasNeuro SinapsisTatiana EspinoAún no hay calificaciones
- Neurofisio Clase 3Documento3 páginasNeurofisio Clase 3Susana LiAún no hay calificaciones
- Sistema nervioso central: Transmisores y receptoresDocumento16 páginasSistema nervioso central: Transmisores y receptoresClaudia MaldonadoAún no hay calificaciones
- SinapsisDocumento26 páginasSinapsisLuis MartinezAún no hay calificaciones
- UntitledDocumento2 páginasUntitledDanna NuñezAún no hay calificaciones
- Expo de FisiologiaDocumento65 páginasExpo de Fisiologiaabi100% (1)
- Mecanismos Celulares Del AprendizajeDocumento4 páginasMecanismos Celulares Del AprendizajeLiliana Bernabe Uribe50% (4)
- Grupo 4Documento24 páginasGrupo 4Luis Gustavo BáezAún no hay calificaciones
- 2.1 ColinergicosDocumento5 páginas2.1 ColinergicosCarola Villanueva MassardoAún no hay calificaciones
- Resumen SINAPSISDocumento10 páginasResumen SINAPSISEsme Monroy0% (1)
- Tema 4. Transmisión SinápticaDocumento9 páginasTema 4. Transmisión SinápticaJose García SegoviaAún no hay calificaciones
- FisiologiaDocumento32 páginasFisiologiawilliamAún no hay calificaciones
- Preguntes FISIODocumento8 páginasPreguntes FISIOmargotAún no hay calificaciones
- CotransmisionDocumento19 páginasCotransmisionGabyta Paladines YangeAún no hay calificaciones
- Sistema LímbicoDocumento5 páginasSistema Límbicotere andersonAún no hay calificaciones
- Memoria y anatomía del hipocampoDocumento7 páginasMemoria y anatomía del hipocampoandhrea rojasAún no hay calificaciones
- Sistema NerviosoDocumento32 páginasSistema Nerviosowskfdw98y7Aún no hay calificaciones
- Clase 3 Fisiología 22 - 04 - 19 PDFDocumento10 páginasClase 3 Fisiología 22 - 04 - 19 PDFTDGAún no hay calificaciones
- Cuestionario S.N.C. Emperatriz IngaDocumento9 páginasCuestionario S.N.C. Emperatriz IngaEmperatrizIngaHidalgoAún no hay calificaciones
- Sinapsis ResumenDocumento9 páginasSinapsis ResumenAniretac Neko Seyer50% (4)
- Asignación SINAPSIS y NEUROTRANSMISORESDocumento7 páginasAsignación SINAPSIS y NEUROTRANSMISORESPaola AlmonteAún no hay calificaciones
- Neurona, Conducción Nerviosa y NeurotransmisoresDocumento7 páginasNeurona, Conducción Nerviosa y NeurotransmisoresRobertoAún no hay calificaciones
- Fisiología 2Documento14 páginasFisiología 2Rocío TempranoAún no hay calificaciones
- Al Final de Los Sentidos 1Documento9 páginasAl Final de Los Sentidos 1Fernanda BarbaAún no hay calificaciones
- Clase 3 Placa NeuromuscularDocumento23 páginasClase 3 Placa NeuromuscularSteve GomezAún no hay calificaciones
- Celula de Merkel 2Documento4 páginasCelula de Merkel 2Estefanía Daza CisnerosAún no hay calificaciones
- Celula de MerkelDocumento8 páginasCelula de MerkelEstefanía Daza CisnerosAún no hay calificaciones
- Tema 4. SinapsisDocumento10 páginasTema 4. SinapsisAlberto LeikeseAún no hay calificaciones
- Sistema límbico y sus estructurasDocumento16 páginasSistema límbico y sus estructurasCARLOS SALVADOR GERONIMO ESPINOAún no hay calificaciones
- Transmisión Sináptica Capitulo 5 TERMINADO.Documento6 páginasTransmisión Sináptica Capitulo 5 TERMINADO.Abril ElisAún no hay calificaciones
- Word Preguntas FisiologiaDocumento3 páginasWord Preguntas Fisiologiasarai villenaAún no hay calificaciones
- La transducción visual: del estímulo a la percepciónDocumento2 páginasLa transducción visual: del estímulo a la percepciónMelissa MatuteAún no hay calificaciones
- Clase Sistema Nervioso Dr. TelloDocumento41 páginasClase Sistema Nervioso Dr. TelloDIEGO ALFONSO FERNANDEZ CALLEAún no hay calificaciones
- Preguntas Tipo Examen FisiologíaDocumento4 páginasPreguntas Tipo Examen FisiologíaArmando Martin LopezAún no hay calificaciones
- Neuroplasticidad (Ficha de Cátedra)Documento6 páginasNeuroplasticidad (Ficha de Cátedra)damianAún no hay calificaciones
- Teoria Delm Musculo CardiacoDocumento12 páginasTeoria Delm Musculo CardiacoAlbert KantAún no hay calificaciones
- AcetilcolinaDocumento2 páginasAcetilcolinaPRISCILA YAMILET JIMENEZ MANZOAún no hay calificaciones
- Sistema NerviosoDocumento3 páginasSistema NerviosoNashely CastilloAún no hay calificaciones
- Capítulo 46 FisioDocumento8 páginasCapítulo 46 Fisioevelyn ramgoAún no hay calificaciones
- Cardiología CTODocumento95 páginasCardiología CTOEfren Penaflores100% (1)
- Las Neuronas y Las Clases de SinapsisDocumento8 páginasLas Neuronas y Las Clases de SinapsisVenus Delta Ticona FloresAún no hay calificaciones
- Resumen Cardiovascular 2 PDFDocumento28 páginasResumen Cardiovascular 2 PDFRuth Maribel López DuarteAún no hay calificaciones
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012De EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012Aún no hay calificaciones
- El cannabis en patologías del sistema nervioso centralDe EverandEl cannabis en patologías del sistema nervioso centralAún no hay calificaciones
- Atención de Salud Mental y AdolescentesDocumento2 páginasAtención de Salud Mental y AdolescentesAngel Stevens Garcia CallaoAún no hay calificaciones
- Manejo Del Paciente PolitraumatizadoDocumento33 páginasManejo Del Paciente PolitraumatizadoAngel Stevens Garcia CallaoAún no hay calificaciones
- Seminario EapDocumento18 páginasSeminario EapAngel Stevens Garcia CallaoAún no hay calificaciones
- Impacto del aislamiento social en la salud mental de adolescentes durante COVID-19Documento8 páginasImpacto del aislamiento social en la salud mental de adolescentes durante COVID-19Angel Stevens Garcia CallaoAún no hay calificaciones
- Sindrome CompartimentalDocumento27 páginasSindrome CompartimentalAngel Stevens Garcia CallaoAún no hay calificaciones
- Control de SangradoDocumento17 páginasControl de SangradoAngel Stevens Garcia CallaoAún no hay calificaciones
- Caso clínico queratitis: diagnóstico y tratamientoDocumento29 páginasCaso clínico queratitis: diagnóstico y tratamientoAngel Stevens Garcia Callao100% (1)
- Libro de ReclamacionesDocumento1 páginaLibro de ReclamacionesAngel Stevens Garcia CallaoAún no hay calificaciones
- GASTROVILLADocumento22 páginasGASTROVILLAAngel Stevens Garcia CallaoAún no hay calificaciones
- $rudw6fo PDFDocumento24 páginas$rudw6fo PDFAngel Stevens Garcia CallaoAún no hay calificaciones
- $rudw6fo PDFDocumento24 páginas$rudw6fo PDFAngel Stevens Garcia CallaoAún no hay calificaciones
- Signosy SintomasDocumento15 páginasSignosy SintomasAngel Stevens Garcia Callao100% (1)
- Grupo 01 - Seminario Enfermedad DiverticularDocumento30 páginasGrupo 01 - Seminario Enfermedad DiverticularAngel Stevens Garcia CallaoAún no hay calificaciones
- Grupo2 Patologia Vesicula y Vias BiliaresDocumento44 páginasGrupo2 Patologia Vesicula y Vias BiliaresAngel Stevens Garcia CallaoAún no hay calificaciones
- $RPYWNUDDocumento36 páginas$RPYWNUDAngel Stevens Garcia CallaoAún no hay calificaciones
- CEIO MetálicoDocumento26 páginasCEIO MetálicoAngel Stevens Garcia CallaoAún no hay calificaciones
- Declaraciones juradas anexosDocumento10 páginasDeclaraciones juradas anexosCésar Carmona BernillaAún no hay calificaciones
- Fractura ExpuestaDocumento32 páginasFractura ExpuestaAngel Stevens Garcia CallaoAún no hay calificaciones
- Insufiencia RespiratoriaDocumento35 páginasInsufiencia RespiratoriaAngel Stevens Garcia CallaoAún no hay calificaciones
- COTIZACIÓNDocumento2 páginasCOTIZACIÓNAngel Stevens Garcia CallaoAún no hay calificaciones
- Prueba ADocumento11 páginasPrueba Aavengersrm 2019Aún no hay calificaciones
- Caso ClinicoDocumento1 páginaCaso ClinicoAngel Stevens Garcia CallaoAún no hay calificaciones
- CervixDocumento14 páginasCervixAngel Stevens Garcia CallaoAún no hay calificaciones
- Dpi ArtrografiaDocumento12 páginasDpi ArtrografiaAngel Stevens Garcia CallaoAún no hay calificaciones
- 2da Parte HCDocumento4 páginas2da Parte HCAngel Stevens Garcia CallaoAún no hay calificaciones
- S35-05 63 - IiiDocumento8 páginasS35-05 63 - IiiAngel Stevens Garcia CallaoAún no hay calificaciones
- Libro ProstataDocumento7 páginasLibro ProstataAlexandra Zapata CórdovaAún no hay calificaciones
- EB04-03 RadiologiaDocumento23 páginasEB04-03 RadiologiaAngel Stevens Garcia CallaoAún no hay calificaciones
- Farmacología GeneralDocumento71 páginasFarmacología GeneralCandela GarciaAún no hay calificaciones
- Farmacologia de Las Sustancias Psicoactivas 2022-23. ALUMNOSDocumento26 páginasFarmacologia de Las Sustancias Psicoactivas 2022-23. ALUMNOSAnalí Sobreyra JuárezAún no hay calificaciones
- Deporte infantil y juvenil: etapas evolutivas en el fútbolDocumento12 páginasDeporte infantil y juvenil: etapas evolutivas en el fútbolLuis Vásquez TelloAún no hay calificaciones
- Guía de Aprendizaje Ciencias NaturalesDocumento13 páginasGuía de Aprendizaje Ciencias NaturalesHernan GomezAún no hay calificaciones
- Factores Que Influencian en El Neurodesarro PDFDocumento67 páginasFactores Que Influencian en El Neurodesarro PDFNora Sedem100% (1)
- Cerebro 1Documento16 páginasCerebro 1claudiaAún no hay calificaciones
- ReporteDocumento9 páginasReportecarlos enrique0% (1)
- Bases Biológicas Del Comportamiento - D - Evaluación 1Documento3 páginasBases Biológicas Del Comportamiento - D - Evaluación 1GUSTAVO SANCHEZ CERVANTESAún no hay calificaciones
- Capítulo 7 FisiológicaDocumento71 páginasCapítulo 7 FisiológicaJavi CordobaAún no hay calificaciones
- Fortalecimiento de La Lectura Con GamificaciónDocumento70 páginasFortalecimiento de La Lectura Con GamificaciónDavid MolinaAún no hay calificaciones
- Investigación: Redes Neuronales ArtificialesDocumento8 páginasInvestigación: Redes Neuronales ArtificialesMateo ChuquimarcaAún no hay calificaciones
- TEMA La Funcion de RelacionDocumento17 páginasTEMA La Funcion de RelacionBrenda Noemi Travezaño MerinoAún no hay calificaciones
- Bases Biologicas Del ComportamientoDocumento28 páginasBases Biologicas Del Comportamientocrrls14adonisAún no hay calificaciones
- Conduccion y Transmision Del Potencial de Accion Dra Oris L de Calvo MSCDocumento21 páginasConduccion y Transmision Del Potencial de Accion Dra Oris L de Calvo MSCMilagros RosasAún no hay calificaciones
- Biofísica Del Sistema NerviosoDocumento30 páginasBiofísica Del Sistema NerviosoNafe Lopez67% (3)
- Tratamiento de Las Drogodependencias Desde Un Enfoque Sistémico PDFDocumento62 páginasTratamiento de Las Drogodependencias Desde Un Enfoque Sistémico PDFEsteysil Molina TomásAún no hay calificaciones
- Anatomía Humana PDFDocumento177 páginasAnatomía Humana PDFRichard Rodriguez GarciaAún no hay calificaciones
- Sistema nervioso: receptores sensitivos y transmisión del dolorDocumento8 páginasSistema nervioso: receptores sensitivos y transmisión del dolorelizabeth alejandraAún no hay calificaciones
- Impulsividad y Agresion Aspectos ClinicosDocumento11 páginasImpulsividad y Agresion Aspectos ClinicosneuropsicojcAún no hay calificaciones
- Sinapsis Dinamica - En.esDocumento13 páginasSinapsis Dinamica - En.essofia.rvvvAún no hay calificaciones
- NeurofisiologíaDocumento95 páginasNeurofisiologíapilarAún no hay calificaciones
- Neralidades Del Sistema NerviosoDocumento71 páginasNeralidades Del Sistema NerviosoMayra Itzel Gonzalez GonzalezAún no hay calificaciones
- 6 Comunicacion CelularDocumento16 páginas6 Comunicacion CelularMarco Arturo Garcia PerezAún no hay calificaciones
- Via Auditiva EferenteDocumento4 páginasVia Auditiva EferenteDaniela LemboAún no hay calificaciones
- Sistema nervioso: generalidades y clasificación anatómica y funcional (38Documento40 páginasSistema nervioso: generalidades y clasificación anatómica y funcional (38José Angel MateoAún no hay calificaciones
- Tipos de neuroplasticidad y mecanismos de producciónDocumento4 páginasTipos de neuroplasticidad y mecanismos de producciónJoseNeiserCastilloArrascue100% (3)
- Las neuronas: estructura y funciónDocumento18 páginasLas neuronas: estructura y funciónDiego A TorrealbaAún no hay calificaciones
- Wuolah Free TP 1 GENERALIDADES DEL SISTDocumento19 páginasWuolah Free TP 1 GENERALIDADES DEL SISTAymara StiebenAún no hay calificaciones
- Fisiología General ¿ProgramaDocumento26 páginasFisiología General ¿Programaalejandra esparza tejedaAún no hay calificaciones