También podría gustarte
- Apostila de Química Analítica QuantitativaDocumento68 páginasApostila de Química Analítica Quantitativaapi-376356989% (9)
- Escala de Responsividade Social SRSDocumento2 páginasEscala de Responsividade Social SRSScribdTranslations100% (4)
- Microbiologia Médica I: Patógenos e Microbioma HumanoDe EverandMicrobiologia Médica I: Patógenos e Microbioma HumanoAún no hay calificaciones
- Banco de Questões (Nutrição e Dietética)Documento2 páginasBanco de Questões (Nutrição e Dietética)Kelyane PortelaAún no hay calificaciones
- 02 - Sistema de Segurança Pública - SlidesDocumento154 páginas02 - Sistema de Segurança Pública - SlidesJanildo Da Silva Arantes Arantes100% (2)
- Apostila de Imunologia (Pratica)Documento100 páginasApostila de Imunologia (Pratica)yurico122Aún no hay calificaciones
- Apostila Tecnologia de BebidasDocumento128 páginasApostila Tecnologia de BebidasAugusto Andrade100% (1)
- Sistema imunológico e CannabisDe EverandSistema imunológico e CannabisCalificación: 5 de 5 estrellas5/5 (1)
- Imuno IIDocumento26 páginasImuno IIJonatan Dos Santos De OliveiraAún no hay calificaciones
- Resposta Inata e AdaptativaDocumento11 páginasResposta Inata e AdaptativaSamara Santos100% (1)
- Ebook ImunidadeDocumento46 páginasEbook Imunidadecarolinafarias1Aún no hay calificaciones
- E Book Sistema Imunologico e Tipos de VacinasDocumento62 páginasE Book Sistema Imunologico e Tipos de VacinasCarolina Morales100% (1)
- Noções Sistema ImunitárioDocumento10 páginasNoções Sistema ImunitárioRaquel AntunesAún no hay calificaciones
- Regeneração e CicatrizaçãoDocumento58 páginasRegeneração e CicatrizaçãoSamuel MorenoAún no hay calificaciones
- Apostila de Imunologia Clínica 2013Documento87 páginasApostila de Imunologia Clínica 2013sergiomolivAún no hay calificaciones
- Farmacopéia Brasileira 5 Ed. Volume 1 - AnvisaDocumento545 páginasFarmacopéia Brasileira 5 Ed. Volume 1 - AnvisaLuizSantos100% (3)
- Documento 2Documento6 páginasDocumento 2Ton CastroAún no hay calificaciones
- IMUNIDADE INATA E INFLAMAÇÃO CALICH-Imunologia 2 Ed - VPDocumento24 páginasIMUNIDADE INATA E INFLAMAÇÃO CALICH-Imunologia 2 Ed - VPJorge Afiune100% (2)
- Imunidade InataDocumento5 páginasImunidade InataisabelaneuroimunologiaAún no hay calificaciones
- Celulas Sistema ImuneDocumento135 páginasCelulas Sistema ImuneMiriam NutricionistaAún no hay calificaciones
- Aula 7 - Infecções Fúngicas e Mecanismos de Defesa No Indivíduo ImunocompetenteDocumento6 páginasAula 7 - Infecções Fúngicas e Mecanismos de Defesa No Indivíduo Imunocompetente9vvzhn8krvAún no hay calificaciones
- Imunidade Inata ResumoDocumento11 páginasImunidade Inata Resumodouglas alvesAún no hay calificaciones
- Hanseniase Avanços e Desafios Colorido - Pdfcap6Documento26 páginasHanseniase Avanços e Desafios Colorido - Pdfcap6Maureen BorgesAún no hay calificaciones
- Células Do Sistema Imune e AnatomiaDocumento19 páginasCélulas Do Sistema Imune e AnatomiaVitoria Pires FelixAún no hay calificaciones
- Imunidade inata e adaptativa no combate ao cancroDocumento25 páginasImunidade inata e adaptativa no combate ao cancroRita FialhoAún no hay calificaciones
- Resumo ImunoDocumento4 páginasResumo ImunoGustavo AbecassisAún no hay calificaciones
- Imunidade Inata e Reconhecimento de PatógenosDocumento22 páginasImunidade Inata e Reconhecimento de PatógenosfabricioAún no hay calificaciones
- Mecanismos Efetores Da Imunidade HumoralDocumento6 páginasMecanismos Efetores Da Imunidade HumoralGuilherme Schmitt de Andrade100% (1)
- Os mecanismos da imunidade inata e a defesa do organismo contra infecçõesDocumento2 páginasOs mecanismos da imunidade inata e a defesa do organismo contra infecçõesNahenna Suiesná Lima Assunção MonteiroAún no hay calificaciones
- Mecanismos Imunológicos Na Relação Parasito-Hospedeiro e Evasão Da Resposta ImuneDocumento9 páginasMecanismos Imunológicos Na Relação Parasito-Hospedeiro e Evasão Da Resposta ImuneGustavo BrazãoAún no hay calificaciones
- Imunidade Inata: Células e Mecanismos de DefesaDocumento4 páginasImunidade Inata: Células e Mecanismos de DefesaÍtalo PamponetAún no hay calificaciones
- Estudo Dirigido - ArianneDocumento5 páginasEstudo Dirigido - ArianneArlete PereiraAún no hay calificaciones
- Mod. II Prob. IVDocumento15 páginasMod. II Prob. IVgerlan rodriguesAún no hay calificaciones
- Aula 2: Reconhecimento dos microorganismos pela imunidade inataDocumento24 páginasAula 2: Reconhecimento dos microorganismos pela imunidade inataCristiane AliceAún no hay calificaciones
- Monócito AbbasDocumento5 páginasMonócito AbbasJordan LimaAún no hay calificaciones
- Imunidade Inata - Cap 2Documento13 páginasImunidade Inata - Cap 2Natália ChiesaAún no hay calificaciones
- M06P01Documento23 páginasM06P01giovannasouzaAún no hay calificaciones
- 7 - Resumo Imunidade InataDocumento3 páginas7 - Resumo Imunidade InataImunoResumo90% (10)
- Introdução ao Sistema ImunológicoDocumento9 páginasIntrodução ao Sistema ImunológicoAndrieli HauschildtAún no hay calificaciones
- Celulas de Desfesa CertoDocumento10 páginasCelulas de Desfesa CertoKeli Brum MaroccoAún no hay calificaciones
- Resumos de ImunologiaDocumento8 páginasResumos de ImunologiaJuliana LimaAún no hay calificaciones
- Defesa Mecanismos 3Documento19 páginasDefesa Mecanismos 3John WayneAún no hay calificaciones
- Seminário MicroDocumento8 páginasSeminário MicroLucas GomesAún no hay calificaciones
- Celulas Do Sistema ImuneDocumento4 páginasCelulas Do Sistema Imunetchelyhilario90Aún no hay calificaciones
- Resumo Imuno Av1Documento3 páginasResumo Imuno Av1Alexya CavalcantiAún no hay calificaciones
- Imunidade a microrganismosDocumento12 páginasImunidade a microrganismosAderlane LaranjeiraAún no hay calificaciones
- Pathophysiology of sepsis - UpToDateDocumento27 páginasPathophysiology of sepsis - UpToDateNetoAún no hay calificaciones
- Moléculas CHP na apresentação de antígenosDocumento2 páginasMoléculas CHP na apresentação de antígenosMichele lagoAún no hay calificaciones
- Imunidade inata contra fungos e protozoáriosDocumento2 páginasImunidade inata contra fungos e protozoáriosFernanda MotaAún no hay calificaciones
- Exercicio Imuno CertoDocumento6 páginasExercicio Imuno CertoThainara SousaAún no hay calificaciones
- Pratica Resposta ImuneDocumento2 páginasPratica Resposta ImuneThaís Sena100% (1)
- Apostila de ImunoDocumento63 páginasApostila de Imunobruna100% (1)
- Resumo Imunidade InataDocumento11 páginasResumo Imunidade InataOmar HugoAún no hay calificaciones
- Imunologia BásicaDocumento106 páginasImunologia BásicaAndressa SantanaAún no hay calificaciones
- Células do sistema imuneDocumento31 páginasCélulas do sistema imunePaulo TeixeiraAún no hay calificaciones
- Sistema ImunológicoDocumento34 páginasSistema ImunológiconazyalenskaAún no hay calificaciones
- Células Do Sistema ImuneDocumento13 páginasCélulas Do Sistema ImuneAnna OliveiraAún no hay calificaciones
- AGD - Estudo Dirigido 31-10-2022Documento3 páginasAGD - Estudo Dirigido 31-10-2022RAFAELA SANTOS FONTANAAún no hay calificaciones
- Ação da imunidade inata em quadros inflamatóriosDocumento6 páginasAção da imunidade inata em quadros inflamatóriosFrancine RicciAún no hay calificaciones
- Imunidade Inata e AdaptativaDocumento13 páginasImunidade Inata e AdaptativaMisael SousaAún no hay calificaciones
- FVS Imunidade InataDocumento59 páginasFVS Imunidade InatarafaellympsAún no hay calificaciones
- Infecção vs InflamaçãoDocumento15 páginasInfecção vs InflamaçãoSilvia Rodrigues100% (1)
- Antibióticos que atuam como antibacterianosDocumento27 páginasAntibióticos que atuam como antibacterianosDébora Martins100% (1)
- Órgãos linfoides primários e secundáriosDocumento6 páginasÓrgãos linfoides primários e secundáriosmarciofsilva1375% (4)
- Inflamação: migração de leucócitos e reconhecimento de micróbiosDocumento8 páginasInflamação: migração de leucócitos e reconhecimento de micróbiosLaleska GabriellaAún no hay calificaciones
- Sistema Imune e Riscos OcupacionaisDocumento22 páginasSistema Imune e Riscos OcupacionaisSaah BatistaAún no hay calificaciones
- Guia de interpretação de exames laboratoriaisDocumento32 páginasGuia de interpretação de exames laboratoriaisKarellineAún no hay calificaciones
- Referencia 5Documento8 páginasReferencia 5Augusto AndradeAún no hay calificaciones
- Desenvolvimento de Embalagens para MedicamentosDocumento41 páginasDesenvolvimento de Embalagens para MedicamentosAugusto AndradeAún no hay calificaciones
- Manual de Vigilância Epidemiológica de Febre Amarela 2004 PDFDocumento69 páginasManual de Vigilância Epidemiológica de Febre Amarela 2004 PDFAugusto AndradeAún no hay calificaciones
- Diretoria Colegiada 44Documento4 páginasDiretoria Colegiada 44Augusto AndradeAún no hay calificaciones
- Referencia 3Documento5 páginasReferencia 3Augusto AndradeAún no hay calificaciones
- Normas Farmacêuticas 2013Documento51 páginasNormas Farmacêuticas 2013Augusto AndradeAún no hay calificaciones
- CartilhaFarmaciaHospitalar 2013 Web4 (10-04-13)Documento60 páginasCartilhaFarmaciaHospitalar 2013 Web4 (10-04-13)Jessica MarcelinoAún no hay calificaciones
- Cromatografia em Camada DelgadaDocumento41 páginasCromatografia em Camada DelgadaAugusto AndradeAún no hay calificaciones
- EletroforeseDocumento40 páginasEletroforeseElaine GaduAún no hay calificaciones
- Bioquimica I CederjDocumento213 páginasBioquimica I Cederjanon_445490458Aún no hay calificaciones
- Livro 01 - Sistema Único de SaúdeDocumento290 páginasLivro 01 - Sistema Único de SaúdehamiltonaspAún no hay calificaciones
- VirologiafiocruzDocumento96 páginasVirologiafiocruzlr0193Aún no hay calificaciones
- (Alcool) Sim Grasielly Regina Andreata 587 69049Documento36 páginas(Alcool) Sim Grasielly Regina Andreata 587 69049Augusto AndradeAún no hay calificaciones
- Contribuição Ao Conhecimento Químico de Plantas Do Nordeste Do BrasilDocumento168 páginasContribuição Ao Conhecimento Químico de Plantas Do Nordeste Do BrasilAugusto AndradeAún no hay calificaciones
- Noções de Saúde e EspiritismoDocumento1 páginaNoções de Saúde e EspiritismoLuxciana RodriguesAún no hay calificaciones
- CAPÍTULO 3 - Sistemas de Informação em Saúde e Vigilância Epidemiológica - MedicinaNETDocumento8 páginasCAPÍTULO 3 - Sistemas de Informação em Saúde e Vigilância Epidemiológica - MedicinaNETFrancisco LeonardoAún no hay calificaciones
- Leitura e Fichamento PsicologiaDocumento4 páginasLeitura e Fichamento PsicologiaAugusto TavaresAún no hay calificaciones
- 01-Laudo Ubs FaculdadeDocumento70 páginas01-Laudo Ubs FaculdadeFERNANDA PINHEIRO DE OLIVEIRAAún no hay calificaciones
- Relatorio EstágioDocumento25 páginasRelatorio EstágioNáthali SantosAún no hay calificaciones
- Fispq - Tinta Spray Colorgin Azul ColonialDocumento13 páginasFispq - Tinta Spray Colorgin Azul Colonialgerson.refraAún no hay calificaciones
- Tecnico Analises Clinicas LaboratorioDocumento9 páginasTecnico Analises Clinicas LaboratoriosabrinabarreteAún no hay calificaciones
- CIRROSE HEPATICA - Consequências da hipertensão portaDocumento5 páginasCIRROSE HEPATICA - Consequências da hipertensão portabismillahjohnAún no hay calificaciones
- Classificação por transferência na Escola Estadual de UmburaninhaDocumento24 páginasClassificação por transferência na Escola Estadual de UmburaninhaAlmirGPAún no hay calificaciones
- O transe mediúnico: conceito, graus e etapasDocumento6 páginasO transe mediúnico: conceito, graus e etapasYara SousaAún no hay calificaciones
- 2 Tendinite PDFDocumento12 páginas2 Tendinite PDFFabio Sales VieiraAún no hay calificaciones
- Aula 06 Farmacocinética IIDocumento23 páginasAula 06 Farmacocinética IIDodocolinaAún no hay calificaciones
- TCC Manual de Aplicação Do LeanHealthcare 21Documento33 páginasTCC Manual de Aplicação Do LeanHealthcare 21GabrielAún no hay calificaciones
- Farmacologia: histórico, conceitos e formas farmacêuticasDocumento41 páginasFarmacologia: histórico, conceitos e formas farmacêuticasAlan SouzaAún no hay calificaciones
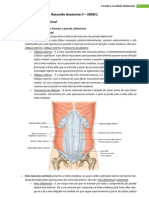
- Resumo de Anatomia - Parede / Cavidade Abdominal e Aparelho DigestorioDocumento47 páginasResumo de Anatomia - Parede / Cavidade Abdominal e Aparelho DigestorioLuiz Felipe100% (17)
- Questionário SWYCDocumento3 páginasQuestionário SWYCLuis Fabricio LojorAún no hay calificaciones
- Orientacao DGS 08 2-03-2011 Carros EmergenciaDocumento11 páginasOrientacao DGS 08 2-03-2011 Carros EmergenciaSofia AraújoAún no hay calificaciones
- Unimed implanta novo marketing estratégicoDocumento10 páginasUnimed implanta novo marketing estratégicoRuth OliveiraAún no hay calificaciones
- Manejo Da Insuficiencia Respiratoria Aguda Guia Pratico 17.11.22Documento39 páginasManejo Da Insuficiencia Respiratoria Aguda Guia Pratico 17.11.22Jonathan Izabel100% (1)
- Aula 2 - Infecção Hospitalar 2016.2Documento17 páginasAula 2 - Infecção Hospitalar 2016.2Jonathan sAún no hay calificaciones
- Medicamento para asma e DPOCDocumento2 páginasMedicamento para asma e DPOCJucemar SilvaAún no hay calificaciones
- Avaliação e Tratamento de FeridasDocumento44 páginasAvaliação e Tratamento de FeridasLuisAún no hay calificaciones
- Aula 01 MNTDocumento30 páginasAula 01 MNTSamuel Belarmino MaunzeAún no hay calificaciones
- Revista Do Observatório Brasil Da Igualdade de GêneroDocumento92 páginasRevista Do Observatório Brasil Da Igualdade de GêneroRayane Lula AndradeAún no hay calificaciones
- Farmácia de ManipulaçãoDocumento6 páginasFarmácia de ManipulaçãoLaura FerreiraAún no hay calificaciones
- Guia Medico - Rede SergipeDocumento28 páginasGuia Medico - Rede SergipeRenato SilveiraAún no hay calificaciones