También podría gustarte
- Hola Soy Tu HigadoDocumento10 páginasHola Soy Tu HigadoshamballaAún no hay calificaciones
- Apuntes Ciclo CelularDocumento5 páginasApuntes Ciclo CelularCristina Sanz100% (1)
- INMUNOLOGÍADocumento5 páginasINMUNOLOGÍAANGELICA MARIA ROJAS MAYORAún no hay calificaciones
- Citología 1 Bach PDFDocumento27 páginasCitología 1 Bach PDFProf. Víctor M. VitoriaAún no hay calificaciones
- Tema 1 (La Célula Procariota) PDFDocumento16 páginasTema 1 (La Célula Procariota) PDFNonde OneneAún no hay calificaciones
- Resumen Fisiologia CelularDocumento8 páginasResumen Fisiologia CelularMario Javier Ganoza Morales100% (1)
- 13) Reticulo EndoplasmicoDocumento27 páginas13) Reticulo EndoplasmicoPerla Gutiérrez RogelAún no hay calificaciones
- Tema 7 Uniones CelularesDocumento6 páginasTema 7 Uniones CelularesDavid Molina LópezAún no hay calificaciones
- Señalización Celular - Matriz ExtracelularDocumento7 páginasSeñalización Celular - Matriz ExtracelularEnzo Guillermo RamirezAún no hay calificaciones
- Bioquimica ApuntesDocumento23 páginasBioquimica ApuntesDeejay JoseAún no hay calificaciones
- Biofisica MuscularDocumento19 páginasBiofisica MuscularJose Marco Muñoz BocanegraAún no hay calificaciones
- Higado UPSJBDocumento27 páginasHigado UPSJBchuwi06Aún no hay calificaciones
- Fisiologia CelularDocumento13 páginasFisiologia CelularJuan Benitto Moreno Rivera100% (1)
- Estructura y Función de La Membrana PlasmáticaDocumento102 páginasEstructura y Función de La Membrana Plasmátical@sm@rinos1100% (1)
- Campos de estudio biologíaDocumento6 páginasCampos de estudio biologíaEmily MoraAún no hay calificaciones
- Genetica Clínica Apuntes CompletosDocumento54 páginasGenetica Clínica Apuntes CompletosfrhasseyAún no hay calificaciones
- Inmunidad AdquiridaDocumento4 páginasInmunidad AdquiridaVicky OñateAún no hay calificaciones
- RibosomasDocumento8 páginasRibosomasAndreaAún no hay calificaciones
- Etología Aplicada Al Enriquecimiento y Bienestar Animal: Tortuga OliváceaDocumento99 páginasEtología Aplicada Al Enriquecimiento y Bienestar Animal: Tortuga OliváceaKarumbéAún no hay calificaciones
- BiofisicaDocumento31 páginasBiofisicaLucia OlmosAún no hay calificaciones
- Retículo Endoplásmico Pi MZVDocumento29 páginasRetículo Endoplásmico Pi MZVCool BoyLVAún no hay calificaciones
- Apuntes Bioquimica Facultad de QuimicaDocumento31 páginasApuntes Bioquimica Facultad de QuimicaLuzAvAún no hay calificaciones
- CICLO DE KREBS FLDocumento32 páginasCICLO DE KREBS FLGabriela CarmonaAún no hay calificaciones
- Líquidos y ElectrolitosDocumento28 páginasLíquidos y ElectrolitosehtoledocAún no hay calificaciones
- Ficha NucleótidosDocumento2 páginasFicha NucleótidosProf. Víctor M. VitoriaAún no hay calificaciones
- Lisosomas y Sus FuncionesDocumento2 páginasLisosomas y Sus FuncionesJose BMAún no hay calificaciones
- Resumen Sobre GenéticaDocumento4 páginasResumen Sobre GenéticaRich Guerrero100% (1)
- Citoesqueleto, ResumenDocumento13 páginasCitoesqueleto, ResumenMKsander GarcíaAún no hay calificaciones
- Cromosmas. Mitosis y MeiosisDocumento14 páginasCromosmas. Mitosis y MeiosisSergio Hernandez100% (1)
- Conducción nerviosaDocumento7 páginasConducción nerviosaAstor GarcìaAún no hay calificaciones
- Fosforilacion OxidativaDocumento19 páginasFosforilacion OxidativaQoqe Catalán Durán100% (1)
- Poríferos, cnidarios y platelmintos: primeros niveles de organización animalDocumento16 páginasPoríferos, cnidarios y platelmintos: primeros niveles de organización animalBiomlio Mijoy NimjiAún no hay calificaciones
- Examen de Biologia MoodleDocumento3 páginasExamen de Biologia MoodleLuzz MargAún no hay calificaciones
- Aparato Reproductor FemeninoDocumento95 páginasAparato Reproductor FemeninolatukiisAún no hay calificaciones
- Cuadro de PunnetDocumento5 páginasCuadro de PunnetMaricela LópezAún no hay calificaciones
- Adhesion CelularDocumento30 páginasAdhesion CelularBeto HdzAún no hay calificaciones
- Tema 1 Célula y Bases de Los Principios InmediatosDocumento19 páginasTema 1 Célula y Bases de Los Principios InmediatosMay Garcia Villalta100% (1)
- Esquema Clasificación y Función de Las ProteínasDocumento3 páginasEsquema Clasificación y Función de Las ProteínasProf. Víctor M. Vitoria100% (1)
- Metabolismo. CH Química, GlicólisisDocumento96 páginasMetabolismo. CH Química, GlicólisisEnrique Avila100% (1)
- Las Tres Leyes de MendelDocumento5 páginasLas Tres Leyes de MendelJhonSonjhuGeronimoCosmeAún no hay calificaciones
- La Relación CelularDocumento5 páginasLa Relación Celularfede320d100% (1)
- El dogma central de la biología molecular: ADN a ARN y proteínasDocumento6 páginasEl dogma central de la biología molecular: ADN a ARN y proteínasRaul Huayanay CarranzaAún no hay calificaciones
- Leccion 55 Los Hemicordados X EnteropneustosDocumento46 páginasLeccion 55 Los Hemicordados X EnteropneustosOmar LariosAún no hay calificaciones
- Canales iónicos y potenciales eléctricos en célulasDocumento88 páginasCanales iónicos y potenciales eléctricos en célulasKiara Velasquez100% (2)
- EstesiologiaDocumento7 páginasEstesiologiaAlberto Castro RosasAún no hay calificaciones
- Estructuras y Organelos de Células EucariotasDocumento16 páginasEstructuras y Organelos de Células EucariotasJossary ButtowskiAún no hay calificaciones
- Guía transporte pasivo células 8° básicoDocumento4 páginasGuía transporte pasivo células 8° básicoCintiaAndreaMaciasAlvaradoAún no hay calificaciones
- Theodor SchwannDocumento7 páginasTheodor SchwannDavid JofreAún no hay calificaciones
- 01 - Sangre - Suero y PlasmaDocumento20 páginas01 - Sangre - Suero y PlasmaInés Carmen PastrianAún no hay calificaciones
- 4-6 Difusión y Transporte Pasivo PDFDocumento5 páginas4-6 Difusión y Transporte Pasivo PDFBrayan Andrės CerónAún no hay calificaciones
- Bomba Sodio Potasio, CotrasporteDocumento6 páginasBomba Sodio Potasio, Cotrasportejefferson martinez mesaAún no hay calificaciones
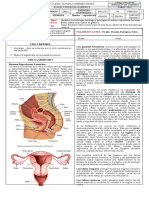
- Guia 11. Reproducción en Humanos - Aparato Reproductor Femenino - 4Documento3 páginasGuia 11. Reproducción en Humanos - Aparato Reproductor Femenino - 4Nicolas Cárdenas maldonadoAún no hay calificaciones
- Seminario Fisiologia Unab 1Documento13 páginasSeminario Fisiologia Unab 1Anonymous ky2jgeaeAún no hay calificaciones
- Guía glicobiología nomenclatura carbohidratosDocumento12 páginasGuía glicobiología nomenclatura carbohidratosSandra R. Manrique HernándezAún no hay calificaciones
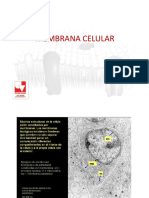
- Membrana Celular y Sistema Endomembranoso PDFDocumento65 páginasMembrana Celular y Sistema Endomembranoso PDFDanielAún no hay calificaciones
- Membrana Print 2.0Documento32 páginasMembrana Print 2.0candelagarciarisekAún no hay calificaciones
- Biologia Apunte DefinitivoDocumento108 páginasBiologia Apunte DefinitivoOlivia CanelaAún no hay calificaciones
- REPASO #2 BIOLOGÍA CyMDocumento28 páginasREPASO #2 BIOLOGÍA CyMMARIA FERNANDA BERROSPI MELGAREJOAún no hay calificaciones
- Tema 2Documento22 páginasTema 2Endika ValladolidAún no hay calificaciones
- Membrana y Transporte PDFDocumento48 páginasMembrana y Transporte PDFMarielos VelásquezAún no hay calificaciones
- boletín 1 semestre 2021 jjvvDocumento2 páginasboletín 1 semestre 2021 jjvvgallardo04Aún no hay calificaciones
- Actividad Asamblea 4 NovDocumento8 páginasActividad Asamblea 4 Novgallardo04Aún no hay calificaciones
- 3.1. INFORME AMBIENTAL TransmarkoDocumento221 páginas3.1. INFORME AMBIENTAL Transmarkogallardo04Aún no hay calificaciones
- ESTUDIO FACTIBILIDAD SANITARIA TransmarkoDocumento97 páginasESTUDIO FACTIBILIDAD SANITARIA Transmarkogallardo04Aún no hay calificaciones
- 1 Modelo Mecano CuanticoDocumento21 páginas1 Modelo Mecano Cuanticogallardo04Aún no hay calificaciones
- Guía 4 Energía en Las Reacciones QuímicasDocumento10 páginasGuía 4 Energía en Las Reacciones Químicasgallardo04Aún no hay calificaciones
- 04 Stem Math WebDocumento24 páginas04 Stem Math Webgallardo04Aún no hay calificaciones
- Preu Prop 2019Documento25 páginasPreu Prop 2019gallardo04Aún no hay calificaciones
- Bifo Berardi - El Tercer IncoscienteDocumento13 páginasBifo Berardi - El Tercer IncoscientePablo Cencig0% (1)
- Guía Formativa A Descubrir Lo Elemental 1° Medio Química NivelaciónDocumento3 páginasGuía Formativa A Descubrir Lo Elemental 1° Medio Química Nivelacióngallardo04Aún no hay calificaciones
- Bases FFCC 2023 2 FSM20102 Iniciativas DepartamentalesDocumento13 páginasBases FFCC 2023 2 FSM20102 Iniciativas Departamentalesgallardo04Aún no hay calificaciones
- Guia 4 Infograma LiquidosDocumento2 páginasGuia 4 Infograma Liquidosgallardo04Aún no hay calificaciones
- Diptico Instructivo - FactoresPsicosociales - Trabajadores-1 56666Documento2 páginasDiptico Instructivo - FactoresPsicosociales - Trabajadores-1 56666diegopascaleAún no hay calificaciones
- Guía 3 Clasificación de Reacciones QuímicasDocumento9 páginasGuía 3 Clasificación de Reacciones Químicasgallardo04Aún no hay calificaciones
- Guía Ejercitación Estructura AtómicaDocumento1 páginaGuía Ejercitación Estructura Atómicagallardo04Aún no hay calificaciones
- Miniensayo 2Documento10 páginasMiniensayo 2gallardo04Aún no hay calificaciones
- 2 Fundamentacion de La ProblematicaDocumento3 páginas2 Fundamentacion de La Problematicagallardo04Aún no hay calificaciones
- Miniensayo MateDocumento9 páginasMiniensayo Mategallardo04Aún no hay calificaciones
- Guia 2 Interpretacion de EtiquetasDocumento2 páginasGuia 2 Interpretacion de Etiquetasgallardo04Aún no hay calificaciones
- 2 Fundamentacion de La ProblematicaDocumento3 páginas2 Fundamentacion de La Problematicagallardo04Aún no hay calificaciones
- Alfabetización científica: conceptos y objetivosDocumento9 páginasAlfabetización científica: conceptos y objetivosjacktroppos100% (2)
- Guia 3 Infograma MasaDocumento2 páginasGuia 3 Infograma Masagallardo04Aún no hay calificaciones
- Enseñar y Aprender Ciencias Algunas Reflexiones. Neus SanmartíDocumento35 páginasEnseñar y Aprender Ciencias Algunas Reflexiones. Neus SanmartíMimi Chile Disfraces100% (6)
- Guia 1 Alimentación y NutriciónDocumento3 páginasGuia 1 Alimentación y Nutricióngallardo04Aún no hay calificaciones
- Orbital EsDocumento1 páginaOrbital Esgallardo04Aún no hay calificaciones
- PlantasDocumento7 páginasPlantasgallardo04Aún no hay calificaciones
- Prueba 1Documento1 páginaPrueba 1gallardo04Aún no hay calificaciones
- Aminoácidos y Péptidos.2017Documento1 páginaAminoácidos y Péptidos.2017gallardo04Aún no hay calificaciones
- Bases Formacion TP (FINAL Nov 2013)Documento104 páginasBases Formacion TP (FINAL Nov 2013)Claudia Andrea Osorio AmesticaAún no hay calificaciones
- BIOENERGÉTICADocumento2 páginasBIOENERGÉTICAgallardo04Aún no hay calificaciones
- Minerales ópticosDocumento13 páginasMinerales ópticosyaku1618100% (1)
- Memoria Practica 3 César Ruiz AlonsoDocumento5 páginasMemoria Practica 3 César Ruiz Alonsocesar ruizAún no hay calificaciones
- Componentes Del Sistema SolarDocumento2 páginasComponentes Del Sistema SolarJessika SotoAún no hay calificaciones
- FLORA APICOLA y NUTRIENTESDocumento4 páginasFLORA APICOLA y NUTRIENTESBenjamín VásquezAún no hay calificaciones
- Prelab 3 - Cinetica de Decoloracion FenolftaleinaDocumento12 páginasPrelab 3 - Cinetica de Decoloracion FenolftaleinaBenjamin Quena CastroAún no hay calificaciones
- Angulos VerticalesDocumento2 páginasAngulos VerticalesMiriam OrmeñoAún no hay calificaciones
- Orígenes coloniales del puerto de BuenaventuraDocumento27 páginasOrígenes coloniales del puerto de BuenaventuraMaría Del Carmen Muñoz Millán100% (1)
- VIGASDocumento40 páginasVIGASEdissonJavierAún no hay calificaciones
- Ud 2 - Valoración InicialDocumento39 páginasUd 2 - Valoración InicialKiddà Și AtâtAún no hay calificaciones
- XCeed InsertDocumento4 páginasXCeed InsertDiego VGAún no hay calificaciones
- Proyecto OlmosDocumento5 páginasProyecto OlmosFrancisco Montoya RiosAún no hay calificaciones
- Quali MicroeconomíaDocumento11 páginasQuali MicroeconomíaMARY LOLY PAREDES MONTALVOAún no hay calificaciones
- ARREGLO GENERAL Preliminar-2020 - RE ANGELUS GABRIEL - L-27750 Mm-ModelDocumento1 páginaARREGLO GENERAL Preliminar-2020 - RE ANGELUS GABRIEL - L-27750 Mm-ModelHéctor MedinaAún no hay calificaciones
- Menú Cocina Zakani Pereira CerritosDocumento23 páginasMenú Cocina Zakani Pereira CerritosMr. MiloxAún no hay calificaciones
- La Ictiofauna de Los Esteros de CamaguanDocumento13 páginasLa Ictiofauna de Los Esteros de Camaguanveliger100Aún no hay calificaciones
- Elaboracion de Pastas, Ravioles y CanelonesDocumento4 páginasElaboracion de Pastas, Ravioles y CanelonesPameAún no hay calificaciones
- Bateria PPT OkDocumento31 páginasBateria PPT OkLuis Alberto Carpio NuñezAún no hay calificaciones
- Método de La AutoobservaciónDocumento4 páginasMétodo de La AutoobservaciónRodrigo Vargas PérezAún no hay calificaciones
- Depresion Postparto Realidad en El Sistema Publico de Atencion de SaludDocumento6 páginasDepresion Postparto Realidad en El Sistema Publico de Atencion de SaludSaray De la TorreAún no hay calificaciones
- Examen FinalDocumento3 páginasExamen FinalNicolasAún no hay calificaciones
- Potencial de la piscicultura en BoliviaDocumento29 páginasPotencial de la piscicultura en BoliviaNen EcoAún no hay calificaciones
- Flip Flop SRDocumento2 páginasFlip Flop SRLeoAún no hay calificaciones
- FichaQuirurgicaCaninaFelinaDocumento3 páginasFichaQuirurgicaCaninaFelinaMauricio Hinojosa JemioAún no hay calificaciones
- McKENNA - El Manjar de Los DiosesDocumento460 páginasMcKENNA - El Manjar de Los DiosesMarisela Amaya VanegasAún no hay calificaciones
- Londres 4 Días 2 Pax 350 Eur/PaxDocumento3 páginasLondres 4 Días 2 Pax 350 Eur/PaxAinhoa Bonilla GomezAún no hay calificaciones
- Pitidiasis Rosada de GibertDocumento6 páginasPitidiasis Rosada de GibertOdel JavierAún no hay calificaciones
- MEDIDASnDEnPESOnYnVOLUMENnnCONVERSIONESn 585f356024ca41aDocumento4 páginasMEDIDASnDEnPESOnYnVOLUMENnnCONVERSIONESn 585f356024ca41aKevinCifuentes100% (1)
- Hamburguesa EspiritualDocumento4 páginasHamburguesa EspiritualJosue Alanoca PomaAún no hay calificaciones
- LoxoscelismoDocumento10 páginasLoxoscelismoVG GabrielAún no hay calificaciones
- Por qué Dios permite el malDocumento5 páginasPor qué Dios permite el malJuan Carlos EberhardtAún no hay calificaciones