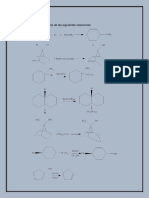
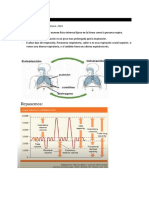
También podría gustarte
- Ciclo de KrebsDocumento6 páginasCiclo de KrebsCesarManuelCastilloCamachoAún no hay calificaciones
- Tablas de Pka PDFDocumento5 páginasTablas de Pka PDFguerreropablo20Aún no hay calificaciones
- Soluciones Ejercicios Formulacion ApuntesDocumento5 páginasSoluciones Ejercicios Formulacion ApuntesNieves Herranz MacíasAún no hay calificaciones
- Sales iónicas: oxisales y haloideasDocumento21 páginasSales iónicas: oxisales y haloideasRaul Milton Aquino BendezuAún no hay calificaciones
- Síntesis de FischerDocumento7 páginasSíntesis de FischerDav AlessiaAún no hay calificaciones
- Capitulos OrganicaDocumento22 páginasCapitulos OrganicaAlonsoAguasZuñigaAún no hay calificaciones
- Síntesis de epóxidos a través de halohidrinas y su apertura mediante nucleófilosDocumento5 páginasSíntesis de epóxidos a través de halohidrinas y su apertura mediante nucleófilosNadia AuzsaAún no hay calificaciones
- Nutrición celular y metabolismoDocumento34 páginasNutrición celular y metabolismoNatasha Jordan100% (2)
- 2.biosintesís de Metabolitos Primarios y Secundarios.2015Documento87 páginas2.biosintesís de Metabolitos Primarios y Secundarios.2015TeffaLo100% (2)
- Ruta del mevalónico para la síntesis de terpenosDocumento13 páginasRuta del mevalónico para la síntesis de terpenosDiplomadoEnTecnicasCromatograficasAún no hay calificaciones
- ComplejosDocumento7 páginasComplejosMaggi Condori GuarachiAún no hay calificaciones
- Reacciones SN y propiedades de nucleofilia y basicidad de alcoholesDocumento21 páginasReacciones SN y propiedades de nucleofilia y basicidad de alcoholesManuel Angelo Rafaile AcostaAún no hay calificaciones
- 7201-14 FISICA MedicionesDocumento31 páginas7201-14 FISICA MedicionesFernando BenottiAún no hay calificaciones
- Introducción A La Nomenclatura de Los Compuestos HeterocíclicosDocumento19 páginasIntroducción A La Nomenclatura de Los Compuestos HeterocíclicosLuis Carlos AndradeAún no hay calificaciones
- MicelasDocumento2 páginasMicelasEduardo EncarnaciónAún no hay calificaciones
- Ejercicios de Afianzamiento 1 Segundo CorteDocumento5 páginasEjercicios de Afianzamiento 1 Segundo CorteMaria SalazarAún no hay calificaciones
- Guía de Mecanismos de Reacción Reacciones de SustituciónDocumento6 páginasGuía de Mecanismos de Reacción Reacciones de SustituciónNaomy Abigail Aguirre Montes de OcaAún no hay calificaciones
- EnzimasDocumento8 páginasEnzimasMARÍA VEGAAún no hay calificaciones
- Separación de Mezclas Por Cristalización y Sublimación.Documento6 páginasSeparación de Mezclas Por Cristalización y Sublimación.paola0% (2)
- Presentacion 6 Cinetica EnzimaticaDocumento46 páginasPresentacion 6 Cinetica EnzimaticaE Clinton LMAún no hay calificaciones
- Tabla LigandosDocumento3 páginasTabla LigandosPaula VelandiaAún no hay calificaciones
- Practica No. 10 SaponificacionDocumento11 páginasPractica No. 10 SaponificacionFernando GarciaAún no hay calificaciones
- Estructuras citoesqueléticas y sus funcionesDocumento6 páginasEstructuras citoesqueléticas y sus funcionesSebastián Rodríguez VelásquezAún no hay calificaciones
- Ácido maleico y fumárico: Preparación y determinación de puntos de fusiónDocumento11 páginasÁcido maleico y fumárico: Preparación y determinación de puntos de fusiónJohan Panez ZevallosAún no hay calificaciones
- Diferenciación de Isómeros de Alcoholes y Propiedades FísicasDocumento16 páginasDiferenciación de Isómeros de Alcoholes y Propiedades FísicasSarmiento DanielAún no hay calificaciones
- FenolesDocumento9 páginasFenolescarlos eduardo vergara lariosAún no hay calificaciones
- Ejercicios de Heterociclos de 5 Miembros Con Un HeteroátomoDocumento8 páginasEjercicios de Heterociclos de 5 Miembros Con Un HeteroátomoIván Narváez AguilarAún no hay calificaciones
- Nomenclatura Aldehidos y Cetonas-ADocumento3 páginasNomenclatura Aldehidos y Cetonas-AAndreas Schilling Morales100% (1)
- Diapositivas Fisicoquimica - TensoactivosDocumento17 páginasDiapositivas Fisicoquimica - Tensoactivosmaicolahumedom84Aún no hay calificaciones
- Introduccion A La Quimica Organica 2016Documento42 páginasIntroduccion A La Quimica Organica 2016Arlethe RodriguezAún no hay calificaciones
- Alcoholes-fenoles-eteresDocumento49 páginasAlcoholes-fenoles-eteresFabiana DazaAún no hay calificaciones
- 6 - Compuestos Oxigenados - Alcoholes, Fenoles y ÉteresDocumento37 páginas6 - Compuestos Oxigenados - Alcoholes, Fenoles y ÉteresShirley LópezAún no hay calificaciones
- Estructura y estabilidad del bencenoDocumento51 páginasEstructura y estabilidad del bencenoeuduardo100% (1)
- Nombre IUPAC de AminoácidosDocumento1 páginaNombre IUPAC de AminoácidosSoledad OjedaAún no hay calificaciones
- Clase 8. Metabolismo EnergéticoDocumento105 páginasClase 8. Metabolismo EnergéticoJuanCaicedo77Aún no hay calificaciones
- La Solvatación Es Un Proceso Que Consiste en La Atracción y Agrupación de Las Moléculas Que Conforman Un DisolventeDocumento7 páginasLa Solvatación Es Un Proceso Que Consiste en La Atracción y Agrupación de Las Moléculas Que Conforman Un DisolventeMaria Jose Barraza CAún no hay calificaciones
- Mecanismos de Las Reacciones OrganicasDocumento24 páginasMecanismos de Las Reacciones OrganicasChule Saul100% (1)
- Taller de ProteínasDocumento9 páginasTaller de ProteínasVALENTINA ANDRADE ANDRADE100% (1)
- Derivados de Acidos CarboxilicosDocumento6 páginasDerivados de Acidos CarboxilicossiriusdarkAún no hay calificaciones
- Comp1 Dismutación Ag - NH3 EZPDocumento2 páginasComp1 Dismutación Ag - NH3 EZPRafael GonzálezAún no hay calificaciones
- Destilacion Por Arrastre de VaporDocumento1 páginaDestilacion Por Arrastre de VaporeduardoAún no hay calificaciones
- Quimica OrganicaDocumento23 páginasQuimica OrganicaOctavio Garcia SotoAún no hay calificaciones
- La Coloración Producida Por El Lugol Se Debe A Que El Yodo Se Introduce Entre Las Espiras de La Molécula de AlmidónDocumento66 páginasLa Coloración Producida Por El Lugol Se Debe A Que El Yodo Se Introduce Entre Las Espiras de La Molécula de Almidónjuan carlosAún no hay calificaciones
- Acidos Carboxilicos y Sus DerivadosDocumento94 páginasAcidos Carboxilicos y Sus DerivadosJr Diego PaxiAún no hay calificaciones
- Cuantificación de Azúcares Reductores y Cromatografía de Carbohidratos en PapelDocumento8 páginasCuantificación de Azúcares Reductores y Cromatografía de Carbohidratos en PapelAngela Liz MuresAún no hay calificaciones
- Ejercicios Química OrgánicaDocumento18 páginasEjercicios Química OrgánicaDaniel Zednanreh ZeugnimodAún no hay calificaciones
- Farmacos Como Mezclas RacemicasDocumento31 páginasFarmacos Como Mezclas RacemicasIrvin Ramirez SalazarAún no hay calificaciones
- Guia 2 Compuestos Oxigenados IDocumento6 páginasGuia 2 Compuestos Oxigenados IGeorgie VegaAún no hay calificaciones
- Nomenclatura de Ácidos CarboxílicosDocumento22 páginasNomenclatura de Ácidos CarboxílicosJorge Poke CervantesAún no hay calificaciones
- Los HeterociclosDocumento21 páginasLos HeterociclosLoré Calderón E100% (1)
- Tablas GlúcidosDocumento6 páginasTablas GlúcidosPablo LordaAún no hay calificaciones
- Resumen Nomenclatura InorganicaDocumento4 páginasResumen Nomenclatura InorganicaAdrian David Bermudez EstupiñanAún no hay calificaciones
- Guias MicrobiologiaDocumento30 páginasGuias MicrobiologiaAndrea100% (1)
- ACIDEZ y BASICIDAD de COMPUESTOS ORGANICOS PDFDocumento13 páginasACIDEZ y BASICIDAD de COMPUESTOS ORGANICOS PDFDaniel Ramírez Ruiz100% (1)
- 5 - Ciclo de KrebsDocumento5 páginas5 - Ciclo de KrebsAngelo Anaya CaponeAún no hay calificaciones
- Ciclo de KrebsDocumento20 páginasCiclo de KrebsCesar CbaAún no hay calificaciones
- Apunte Ciclo Acido CitricoDocumento16 páginasApunte Ciclo Acido CitricoIdelberto BenitezAún no hay calificaciones
- Ciclo de KrepsDocumento12 páginasCiclo de KrepsChapis CtzAún no hay calificaciones
- Ciclo de KrebsDocumento15 páginasCiclo de KrebsMiguel Angel Rodas Herrera100% (2)
- Ciclo de KrebsDocumento8 páginasCiclo de KrebsFabian Herrera MarínAún no hay calificaciones
- Año Del Bicentenario Del PerúDocumento11 páginasAño Del Bicentenario Del PerúCesarManuelCastilloCamachoAún no hay calificaciones
- Tarea Control de La ErosiónDocumento7 páginasTarea Control de La ErosiónCesarManuelCastilloCamachoAún no hay calificaciones
- Tesis Moringa Forrajera Invest IIDocumento17 páginasTesis Moringa Forrajera Invest IICesarManuelCastilloCamachoAún no hay calificaciones
- Cultivo Caña de Azucar PDFDocumento37 páginasCultivo Caña de Azucar PDFCesarManuelCastilloCamachoAún no hay calificaciones
- Fisiología de cultivos I: Fotoperiodismo y fotomorfogénesisDocumento65 páginasFisiología de cultivos I: Fotoperiodismo y fotomorfogénesisCesarManuelCastilloCamachoAún no hay calificaciones
- Alvarado Huaman Leonel EduardoDocumento74 páginasAlvarado Huaman Leonel EduardoCesarManuelCastilloCamachoAún no hay calificaciones
- Producto AcreditableDocumento26 páginasProducto AcreditableCesarManuelCastilloCamachoAún no hay calificaciones
- Clase P CONTROL P Semana 1 PDFDocumento16 páginasClase P CONTROL P Semana 1 PDFCesarManuelCastilloCamachoAún no hay calificaciones
- Tarea #1 de Administracion y GerenciaDocumento4 páginasTarea #1 de Administracion y GerenciaCesarManuelCastilloCamachoAún no hay calificaciones
- Coevolución Planta-InsectoDocumento11 páginasCoevolución Planta-InsectoMaria Delfina Almeyda100% (1)
- Sem 1A.Coevolución e Interacciones Plantas e InsectosDocumento17 páginasSem 1A.Coevolución e Interacciones Plantas e InsectosCesarManuelCastilloCamachoAún no hay calificaciones
- Conceptos básicos de genética: fenotipo, genotipo, cromosomas, genes y másDocumento5 páginasConceptos básicos de genética: fenotipo, genotipo, cromosomas, genes y másCesarManuelCastilloCamachoAún no hay calificaciones
- Deficiencia de NutrientesDocumento10 páginasDeficiencia de NutrientesCesarManuelCastilloCamachoAún no hay calificaciones
- Respiracion VegetalDocumento7 páginasRespiracion VegetalCesarManuelCastilloCamachoAún no hay calificaciones
- Cultivos HidropónicosDocumento20 páginasCultivos HidropónicosCesarManuelCastilloCamachoAún no hay calificaciones
- Codigo GeneticoDocumento8 páginasCodigo GeneticoCesarManuelCastilloCamachoAún no hay calificaciones
- 15 FloraciónDocumento8 páginas15 FloraciónCesarManuelCastilloCamachoAún no hay calificaciones
- GerminacionDocumento6 páginasGerminacionCesarManuelCastilloCamachoAún no hay calificaciones
- Clorofila A y BDocumento5 páginasClorofila A y BCesarManuelCastilloCamachoAún no hay calificaciones
- Meristemas plantas crecimientoDocumento16 páginasMeristemas plantas crecimientoCesarManuelCastilloCamachoAún no hay calificaciones
- Deficiencia de NutrientesDocumento10 páginasDeficiencia de NutrientesCesarManuelCastilloCamachoAún no hay calificaciones
- Respiracion VegetalDocumento7 páginasRespiracion VegetalCesarManuelCastilloCamachoAún no hay calificaciones
- OrgánuloDocumento7 páginasOrgánuloCesarManuelCastilloCamachoAún no hay calificaciones
- Codigo GeneticoDocumento8 páginasCodigo GeneticoCesarManuelCastilloCamachoAún no hay calificaciones
- OrgánuloDocumento7 páginasOrgánuloCesarManuelCastilloCamachoAún no hay calificaciones
- La mitocondria: orgánulo clave en la respiración celularDocumento4 páginasLa mitocondria: orgánulo clave en la respiración celularCesarManuelCastilloCamachoAún no hay calificaciones
- PigmentosDocumento3 páginasPigmentosCesarManuelCastilloCamachoAún no hay calificaciones
- VernalizacionDocumento7 páginasVernalizacionCesarManuelCastilloCamachoAún no hay calificaciones
- Meristemas plantas crecimientoDocumento16 páginasMeristemas plantas crecimientoCesarManuelCastilloCamachoAún no hay calificaciones
- Plastidios y CloroplastoDocumento9 páginasPlastidios y CloroplastoCesarManuelCastilloCamachoAún no hay calificaciones
- Manifestaciones pulmonaresDocumento1 páginaManifestaciones pulmonaresYolanda Zepeda GonzalezAún no hay calificaciones
- Respiración: en Esta Parte Del Examen Físico Interesa Fijarse en La Forma Como La Persona RespiraDocumento38 páginasRespiración: en Esta Parte Del Examen Físico Interesa Fijarse en La Forma Como La Persona RespiralucasAún no hay calificaciones
- Sistema RespiratorioDocumento4 páginasSistema RespiratorioEmmy MixaAún no hay calificaciones
- Tabla 2-1. Parámetros Gasométricos en La Sangre Arterial A - Medicina InternaDocumento1 páginaTabla 2-1. Parámetros Gasométricos en La Sangre Arterial A - Medicina Internasteve edwinAún no hay calificaciones
- Anatomía Del PulmonDocumento3 páginasAnatomía Del PulmonDavid TovarAún no hay calificaciones
- Técnica de Respiracion DiafragmáticaDocumento8 páginasTécnica de Respiracion DiafragmáticaRodrigo OlivaresAún no hay calificaciones
- Presentación VMNIDocumento25 páginasPresentación VMNIssasianAún no hay calificaciones
- Taller Sobre Respiración Celular y FotosintesisDocumento5 páginasTaller Sobre Respiración Celular y FotosintesisMartinAlonsoBuelvasGarrido100% (1)
- MHK Hoberman Sphere 2021 - SpanishDocumento1 páginaMHK Hoberman Sphere 2021 - SpanishNathalie CharlotteAún no hay calificaciones
- Aparato Respiratorio 2022Documento34 páginasAparato Respiratorio 2022Angelo Matteo Zandonai BonaniAún no hay calificaciones
- Practica 05-Fisiología - Peña Castillo LucianaDocumento12 páginasPractica 05-Fisiología - Peña Castillo LucianaLUCIANA FERNANDA PE�A CASTILLOAún no hay calificaciones
- ÑDocumento3 páginasÑANDRE ALEXANDER QUINTAL RUBIOAún no hay calificaciones
- Algoritmos control ventiladores COVID-19Documento7 páginasAlgoritmos control ventiladores COVID-19Alejandra FuenmayorAún no hay calificaciones
- 022 Med Ventilador Mecanico DR Pablo MattosDocumento5 páginas022 Med Ventilador Mecanico DR Pablo MattosWarren Pereyra OrozcoAún no hay calificaciones
- Taller de La RespiraciónDocumento8 páginasTaller de La RespiraciónNini Duarte100% (1)
- Ejercicios Sobre Respiración CelularDocumento4 páginasEjercicios Sobre Respiración CelularanaisAún no hay calificaciones
- 3 PEEP DEsmery PDFDocumento29 páginas3 PEEP DEsmery PDFMaría Alejandra Cerda JanaAún no hay calificaciones
- Sistema RespiratorioDocumento5 páginasSistema RespiratorioByronDanielAún no hay calificaciones
- Respiración Externa y Respiración InternaDocumento30 páginasRespiración Externa y Respiración InternaLihoy KamAún no hay calificaciones
- Clase 7 Sistema RespiratorioDocumento57 páginasClase 7 Sistema RespiratorioalfredoAún no hay calificaciones
- Insuficiencia RespiratoriaDocumento10 páginasInsuficiencia RespiratoriaMica GonzalezAún no hay calificaciones
- Fisiología de La Respiración para Estudiantes de EnfermeriaDocumento21 páginasFisiología de La Respiración para Estudiantes de EnfermeriaJudith Asuncion Lopez Sanchez100% (2)
- 6.-SX PleuropulmonaresDocumento29 páginas6.-SX PleuropulmonaresAlverick MineAún no hay calificaciones
- El Sistema Respiratorio (Anatomía)Documento3 páginasEl Sistema Respiratorio (Anatomía)Alicia GarcíaAún no hay calificaciones
- Anatomía FaringeDocumento13 páginasAnatomía FaringeAlexaB.MendozaAún no hay calificaciones
- Taller No 2 (Respiración Animales) - Biología (Ciclo 3°)Documento3 páginasTaller No 2 (Respiración Animales) - Biología (Ciclo 3°)esteban hernandezAún no hay calificaciones
- SISTEMA - PPTX RespiratorioDocumento10 páginasSISTEMA - PPTX Respiratorioclaudio maldonadoAún no hay calificaciones
- Infografia 3 - RiquelmeDocumento1 páginaInfografia 3 - RiquelmeAlex Javier Riquelme ArévaloAún no hay calificaciones
- Cadena Respiratoria CuevasDocumento3 páginasCadena Respiratoria CuevasLiriany CuevasAún no hay calificaciones
- Sistema Respiratorio y Sistema MuscularDocumento15 páginasSistema Respiratorio y Sistema MuscularEYRA LOPEZAún no hay calificaciones