También podría gustarte
- The Shape of Life: Genes, Development, and the Evolution of Animal FormDe EverandThe Shape of Life: Genes, Development, and the Evolution of Animal FormAún no hay calificaciones
- LectDocumento56 páginasLectSebna NoushadAún no hay calificaciones
- Halanych Annurev - Ecolsys.35.112202.130124 1Documento32 páginasHalanych Annurev - Ecolsys.35.112202.130124 1Adriana MendonçaAún no hay calificaciones
- Primate Evolution: Evidence From The Fossil Record, Comparative Morphology, and Molecular BiologyDocumento16 páginasPrimate Evolution: Evidence From The Fossil Record, Comparative Morphology, and Molecular BiologyWindasari Putri SeptarinaAún no hay calificaciones
- Pgen 1001342Documento18 páginasPgen 1001342Camiz Rmirex ValenciaAún no hay calificaciones
- Evolution and Origin of BiodiversityDocumento13 páginasEvolution and Origin of Biodiversitykharry8davidAún no hay calificaciones
- Cnidarian Milestones in Metazoan Evolution: Ferdinando Boero, Bernd Schierwater, and Stefano PirainoDocumento8 páginasCnidarian Milestones in Metazoan Evolution: Ferdinando Boero, Bernd Schierwater, and Stefano Pirainovltg_35643Aún no hay calificaciones
- Molecular Evolution and PhylogeneticsDocumento16 páginasMolecular Evolution and PhylogeneticsHarsh Vardhan Singh ShekhawatAún no hay calificaciones
- 2 Quarter: Animal Diversity: Learning ObjectivesDocumento11 páginas2 Quarter: Animal Diversity: Learning ObjectivesJella GodoyAún no hay calificaciones
- Genomes 2 Cap16Documento34 páginasGenomes 2 Cap16Angel CarmonaAún no hay calificaciones
- Diversity of Microbial WorldDocumento16 páginasDiversity of Microbial WorldAfrin IbrahimAún no hay calificaciones
- Joram Case StudyDocumento15 páginasJoram Case StudyHoney Bee S. PlatolonAún no hay calificaciones
- UNIT 8 The Paths Traversed by LifeDocumento5 páginasUNIT 8 The Paths Traversed by LifeSindhu SajithAún no hay calificaciones
- Evidences of EvolutionDocumento5 páginasEvidences of EvolutionAnthony CreationAún no hay calificaciones
- Molecular Biology - 12 TESLADocumento9 páginasMolecular Biology - 12 TESLAAlleah Angelica CañoAún no hay calificaciones
- Genbio Mod 2Documento10 páginasGenbio Mod 2ScionAún no hay calificaciones
- Molecular Population Genetics and Evolution - Masatoshi NeiDocumento290 páginasMolecular Population Genetics and Evolution - Masatoshi NeiGeorge Sebastian Antony100% (2)
- Lesson 5 7 Zoology Learning ModuleDocumento38 páginasLesson 5 7 Zoology Learning ModuleAnde Falcone AlbiorAún no hay calificaciones
- Taxonomy and Phylogeny Animal ClassificationDocumento10 páginasTaxonomy and Phylogeny Animal ClassificationTapan Kumar PalAún no hay calificaciones
- Chapter 2 - Evolution of Nervous Systems and Brains (Roth and Dicke) - Book Neurosciences, From Molecule To BehaviorDocumento28 páginasChapter 2 - Evolution of Nervous Systems and Brains (Roth and Dicke) - Book Neurosciences, From Molecule To BehaviorLilian Cerri MazzaAún no hay calificaciones
- The Possibility of Impossible Cultures: HorizonsDocumento7 páginasThe Possibility of Impossible Cultures: HorizonsgeraldAún no hay calificaciones
- Fink The Conceptual Relationship Between Ontogeny and Phylogeny 1982Documento11 páginasFink The Conceptual Relationship Between Ontogeny and Phylogeny 1982JoshuaAún no hay calificaciones
- Reed2017 - Model Organism ConsiderationsDocumento8 páginasReed2017 - Model Organism ConsiderationsIonascu AdrianAún no hay calificaciones
- 2012 - Human Brain Evolution From Gene Discovery To Phenotype DiscoveryDocumento8 páginas2012 - Human Brain Evolution From Gene Discovery To Phenotype DiscoveryMauricio RíosAún no hay calificaciones
- Evidence of EvolutionDocumento10 páginasEvidence of Evolutionr1meshb1lla1969Aún no hay calificaciones
- Miller/Tupper: Zoology 11e Instructor's Manual: Chapter SummaryDocumento3 páginasMiller/Tupper: Zoology 11e Instructor's Manual: Chapter SummaryCaesar Franz RuizAún no hay calificaciones
- An Introduction To ZoologyDocumento13 páginasAn Introduction To ZoologyShafiq KhanAún no hay calificaciones
- General and Comparative Endocrinology: Melissa A. Graham, Ryan L. Earley, John A. Baker, Susan A. FosterDocumento9 páginasGeneral and Comparative Endocrinology: Melissa A. Graham, Ryan L. Earley, John A. Baker, Susan A. FosterGeraldine Villamil JiménezAún no hay calificaciones
- Chick EmbryologyDocumento55 páginasChick EmbryologySushma MishraAún no hay calificaciones
- An Introduction To ZoologyDocumento11 páginasAn Introduction To ZoologySameh AbdelghanyAún no hay calificaciones
- Van - Valen - Homology and CausesDocumento8 páginasVan - Valen - Homology and CausesJulio Torres100% (1)
- Group 6-3ebio: Assoc Prof. Loida R. Medina, PHD, RMT, RmicroDocumento4 páginasGroup 6-3ebio: Assoc Prof. Loida R. Medina, PHD, RMT, RmicroAnne Olfato ParojinogAún no hay calificaciones
- 1635 (2005) Paul B. Eckburg: Science Et AlDocumento5 páginas1635 (2005) Paul B. Eckburg: Science Et AlArizha IrmaAún no hay calificaciones
- Discuss TaxonomyDocumento4 páginasDiscuss TaxonomyAlyssa AlbertoAún no hay calificaciones
- Evolution of ManDocumento2 páginasEvolution of Manxela_g18Aún no hay calificaciones
- 7201 Et ET 07Documento15 páginas7201 Et ET 07Karan veerAún no hay calificaciones
- Seven Types of Pleiotropy: Jonathan HodgkinDocumento5 páginasSeven Types of Pleiotropy: Jonathan HodgkinAriyo 'Jhon Jhay' IbukunAún no hay calificaciones
- Chapter 20 Phylogeny (AP Biology Notes)Documento5 páginasChapter 20 Phylogeny (AP Biology Notes)LeAnnAún no hay calificaciones
- Module 8 - Diversity of Microbial World: Lecture 1-Microbial Taxonomy and PhylogenyDocumento27 páginasModule 8 - Diversity of Microbial World: Lecture 1-Microbial Taxonomy and PhylogenyDr. P.V.Kamala KumariAún no hay calificaciones
- Activity 1 3Documento4 páginasActivity 1 3Nicole Shane PampangaAún no hay calificaciones
- Activity 1 3Documento4 páginasActivity 1 3Nicole Shane PampangaAún no hay calificaciones
- Science and Justice: Lauren P. Chun, Marcus J. Miguel, Emily N. Junkins, Shari L. Forbes, David O. CarterDocumento8 páginasScience and Justice: Lauren P. Chun, Marcus J. Miguel, Emily N. Junkins, Shari L. Forbes, David O. CarterMiguel NadalAún no hay calificaciones
- Evidence of EvolutionDocumento10 páginasEvidence of EvolutionshararohitAún no hay calificaciones
- Comparative Analysis Reveals Epigenomic Evolution Related To Species Traits and Genomic Imprinting in MammalsDocumento67 páginasComparative Analysis Reveals Epigenomic Evolution Related To Species Traits and Genomic Imprinting in Mammals廖晨延Aún no hay calificaciones
- Chapter 26 Study GuideDocumento4 páginasChapter 26 Study GuideBen ReebsAún no hay calificaciones
- Evidences and Bases of EvolutionDocumento21 páginasEvidences and Bases of EvolutionArlynKaren VargasAún no hay calificaciones
- Epigenetics in Teleost Fish - From Molecular Mechanisms To Physiological Phenotypes PDFDocumento35 páginasEpigenetics in Teleost Fish - From Molecular Mechanisms To Physiological Phenotypes PDFTiago TorresAún no hay calificaciones
- Module 13Documento13 páginasModule 13John Bernard VeloriaAún no hay calificaciones
- A Great-Ape View of The Gut MicrobiomeDocumento12 páginasA Great-Ape View of The Gut MicrobiomeHaru SahaAún no hay calificaciones
- Types of Evidence and Procedures That Can Be Used To Establish Evolutionary RelationshipsDocumento20 páginasTypes of Evidence and Procedures That Can Be Used To Establish Evolutionary RelationshipsagpgenabeAún no hay calificaciones
- Unit 1-Change Over Time: InvestigationDocumento5 páginasUnit 1-Change Over Time: InvestigationGnaWYrreJAún no hay calificaciones
- HepatitisDocumento45 páginasHepatitisReza MajidiAún no hay calificaciones
- Emerson Et Al 1988Documento12 páginasEmerson Et Al 1988Sebastián BarrionuevoAún no hay calificaciones
- Epigenetics and Animal Behavior: January 2013Documento26 páginasEpigenetics and Animal Behavior: January 2013Farhad BadshahAún no hay calificaciones
- Role of Cytoplasm in Cell DifferentiationDocumento7 páginasRole of Cytoplasm in Cell DifferentiationJ LALAún no hay calificaciones
- Mini Review ArticleDocumento7 páginasMini Review ArticleEsabel LlanoAún no hay calificaciones
- Diversity of Life (II)Documento176 páginasDiversity of Life (II)tariAún no hay calificaciones
- Kahn Evidence of EvolutionDocumento13 páginasKahn Evidence of EvolutionReychille AbianAún no hay calificaciones
- Listening Test #9Documento3 páginasListening Test #9personalmail_20011078Aún no hay calificaciones
- Austrolebias RevDocumento162 páginasAustrolebias RevFelipe BuenoAún no hay calificaciones
- Lecture 2 - HistologyDocumento7 páginasLecture 2 - HistologyMaddie HallAún no hay calificaciones
- Muscles of The BackDocumento7 páginasMuscles of The BackDon RaulAún no hay calificaciones
- Powerpoint Lectures For: Biology, Seventh EditionDocumento124 páginasPowerpoint Lectures For: Biology, Seventh Editionxo_simpledream100% (1)
- Circulation + Composition of Blood + Vertebrate Heart + Invertebrate Heart + Blood VesselsDocumento121 páginasCirculation + Composition of Blood + Vertebrate Heart + Invertebrate Heart + Blood VesselsReina RamirezAún no hay calificaciones
- 2ant 2Documento3 páginas2ant 2John Robb CorreaAún no hay calificaciones
- The Diagram Shows A Mother Fox and Her CubsDocumento11 páginasThe Diagram Shows A Mother Fox and Her CubsTansra KaurAún no hay calificaciones
- Virology ManualDocumento28 páginasVirology ManualDadan RamadhanAún no hay calificaciones
- MicrosporogenesisDocumento2 páginasMicrosporogenesisYolandhea Firdanasari FirdausAún no hay calificaciones
- Animal Structure Ruminant and MonogastricDocumento23 páginasAnimal Structure Ruminant and MonogastricQuinnee VallejosAún no hay calificaciones
- Animal KingdomDocumento67 páginasAnimal KingdomNaveen Krishna M.KAún no hay calificaciones
- Unit 1 Living Things Natural Science 3ºlast PDFDocumento34 páginasUnit 1 Living Things Natural Science 3ºlast PDFJosefa Jerez0% (1)
- The Impact of Insect Pests On Cowpea Production in NigeriaDocumento35 páginasThe Impact of Insect Pests On Cowpea Production in NigeriaOmowunmi A-TAún no hay calificaciones
- ALPHABET-activity Worksheets For KinderDocumento84 páginasALPHABET-activity Worksheets For KinderLeah Paulyn Bascos Bergonio-LozadaAún no hay calificaciones
- Kottelat 2013 Suplemen - s27Documento667 páginasKottelat 2013 Suplemen - s27Lord GeonagaAún no hay calificaciones
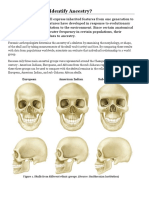
- Activity: Can You Identify Ancestry?: European American Indian Sub-Saharan AfricanDocumento5 páginasActivity: Can You Identify Ancestry?: European American Indian Sub-Saharan AfricanpoovenmoonoosamyAún no hay calificaciones
- 5 - Oral Mucosa and Salivary Glands (Mahmoud Bakr)Documento115 páginas5 - Oral Mucosa and Salivary Glands (Mahmoud Bakr)MobarobberAún no hay calificaciones
- JKSSB Class IV 27 Feb Paper JK Latest InfoDocumento12 páginasJKSSB Class IV 27 Feb Paper JK Latest InfoIdrees AhmedAún no hay calificaciones
- B.SC 1st Yr Syl. 101 & 102-1Documento3 páginasB.SC 1st Yr Syl. 101 & 102-1Anwita JhaAún no hay calificaciones
- Jolly Phonics Printable BookDocumento35 páginasJolly Phonics Printable Bookmeemoo shiekhAún no hay calificaciones
- BIODIVERSITI f2 2017Documento54 páginasBIODIVERSITI f2 2017farah6214100% (1)
- Introduction To The HelminthsDocumento7 páginasIntroduction To The HelminthsmuftizamanAún no hay calificaciones
- Excretory Products and Their EliminationDocumento9 páginasExcretory Products and Their EliminationBiju MylachalAún no hay calificaciones
- NURSAKINAH NAJWAH BT SUHAIMI (Dissection and Observation of Plant Reproductive Organs)Documento9 páginasNURSAKINAH NAJWAH BT SUHAIMI (Dissection and Observation of Plant Reproductive Organs)Nursakinah NajwahAún no hay calificaciones
- K.14 Histology of Nasopharynx & PleuraDocumento26 páginasK.14 Histology of Nasopharynx & Pleuraenri0% (1)
- Basic Sight WordsDocumento9 páginasBasic Sight WordsCasandra PontemayorAún no hay calificaciones
- Overview of Hominin EvolutionDocumento5 páginasOverview of Hominin EvolutionJaviera GibertoAún no hay calificaciones
- Seçmeli Ingilizce Kısaltılmış2021Documento4 páginasSeçmeli Ingilizce Kısaltılmış2021yaseminAún no hay calificaciones
- Propose A SolutionDocumento21 páginasPropose A SolutionPhước NguyễnAún no hay calificaciones