También podría gustarte
- Historia Clinica Juana CH QuemadDocumento13 páginasHistoria Clinica Juana CH QuemadMiguel Roque-Tuero100% (1)
- 10° BiolDocumento5 páginas10° Biolsergey ferrer100% (1)
- Dormancia de PropagulosDocumento2 páginasDormancia de PropagulosEVELYN VALDES CUELLOAún no hay calificaciones
- El Vicepresidente de YoongiDocumento137 páginasEl Vicepresidente de YoongiYndira TovarAún no hay calificaciones
- Sistema DigestivoDocumento38 páginasSistema DigestivoLludith Lazo Tananta100% (1)
- Fisiologia Columna Lumbar UTDocumento49 páginasFisiologia Columna Lumbar UTMiguel Borda0% (1)
- Los Genios No NacenDocumento5 páginasLos Genios No NacenEdwin Gonzalito100% (1)
- Guía básica de términos en quechuaDocumento9 páginasGuía básica de términos en quechuaMuni HvcaAún no hay calificaciones
- Infografia MitocondriaDocumento1 páginaInfografia MitocondriaCesar osvaldo Sanchez RamirezAún no hay calificaciones
- Spanish User Manual-Medonic CA 530 620Documento122 páginasSpanish User Manual-Medonic CA 530 620Patricia MendozaAún no hay calificaciones
- Aminoacidos No EsencialesDocumento1 páginaAminoacidos No EsencialesYOSMERY SANCHEZAún no hay calificaciones
- BIOMOLÉCULASDocumento4 páginasBIOMOLÉCULASMarco Eduardo RodasAún no hay calificaciones
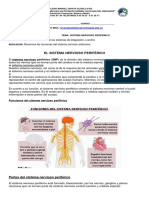
- Sistema nervioso periférico: Estructura y funciones claveDocumento8 páginasSistema nervioso periférico: Estructura y funciones claveHangy Martínez ArrietaAún no hay calificaciones
- Guia INSTITUCIÓN EDUCATIVA VILLA MARGARITADocumento6 páginasGuia INSTITUCIÓN EDUCATIVA VILLA MARGARITADaniiaa BerrocalAún no hay calificaciones
- Bioquimica BasicaDocumento17 páginasBioquimica BasicaLucia VillacrizAún no hay calificaciones
- TRX CircuitoDocumento6 páginasTRX CircuitoPrudenAún no hay calificaciones
- BIBLEOTECADocumento8 páginasBIBLEOTECAROGA CONSTRUCCIONES NAYARITASAún no hay calificaciones
- Anatomía del sistema urogenital masculino y femeninoDocumento39 páginasAnatomía del sistema urogenital masculino y femeninoCamila Pereira BambirraAún no hay calificaciones
- Shiatsu y Las Cinco Etapas de La Transformación de La EnergíaDocumento3 páginasShiatsu y Las Cinco Etapas de La Transformación de La EnergíaGastón Ismael GonzalezAún no hay calificaciones
- Síndrome Pos RCPDocumento88 páginasSíndrome Pos RCPDavid Gresky Avellaneda Talledo100% (1)
- Ciclooxigenasas COXDocumento4 páginasCiclooxigenasas COXLuis Castro Xtrm100% (1)
- Músculos de La CabezaDocumento3 páginasMúsculos de La CabezaCádiz De la hozAún no hay calificaciones
- Placenta y membranas fetales: estructura y funciónDocumento4 páginasPlacenta y membranas fetales: estructura y funciónDiana CabargaAún no hay calificaciones
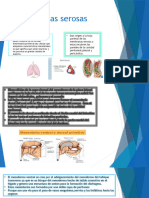
- Membranas SerosasDocumento7 páginasMembranas SerosasDoge LELAún no hay calificaciones
- Toxemia Del EmbarazoDocumento19 páginasToxemia Del EmbarazoMichell Tabora100% (1)
- Una Pasion en El DesiertoDocumento8 páginasUna Pasion en El DesiertoyolotlbAún no hay calificaciones
- Fisiologia CelularDocumento13 páginasFisiologia CelularJuan Benitto Moreno Rivera100% (1)
- Sistema Urinario-2Documento52 páginasSistema Urinario-2Samantha ZuñigaAún no hay calificaciones
- Descubrimiento de La InsulinaDocumento6 páginasDescubrimiento de La Insulinaazucenamart87Aún no hay calificaciones
- Quemosis PDFDocumento3 páginasQuemosis PDFTJHORNA100% (1)