También podría gustarte
- Shoe Dog: A Memoir by the Creator of NikeDe EverandShoe Dog: A Memoir by the Creator of NikeCalificación: 4.5 de 5 estrellas4.5/5 (537)
- Improve Problem Solving With Critical ThinkingDocumento1 páginaImprove Problem Solving With Critical ThinkingIsmael Vara CarvalhoAún no hay calificaciones
- The Yellow House: A Memoir (2019 National Book Award Winner)De EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Calificación: 4 de 5 estrellas4/5 (98)
- Neuromuscular Responses To Isometric Concentric and Eccentric Contractions of The Knee Extensors at The Same Torque Time IntegralDocumento15 páginasNeuromuscular Responses To Isometric Concentric and Eccentric Contractions of The Knee Extensors at The Same Torque Time IntegralIsmael Vara CarvalhoAún no hay calificaciones
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe EverandNever Split the Difference: Negotiating As If Your Life Depended On ItCalificación: 4.5 de 5 estrellas4.5/5 (838)
- História AngiografiaDocumento13 páginasHistória AngiografiaIsmael Vara CarvalhoAún no hay calificaciones
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceCalificación: 4 de 5 estrellas4/5 (890)
- Handbook of Osteopathic TechniqueDocumento289 páginasHandbook of Osteopathic Techniquemchus1100% (14)
- Grit: The Power of Passion and PerseveranceDe EverandGrit: The Power of Passion and PerseveranceCalificación: 4 de 5 estrellas4/5 (587)
- 2006 Management of Peripheral Neuropathic Pain. Integrating Neurobiology, Neurodynamics and Clinical EvidenceDocumento14 páginas2006 Management of Peripheral Neuropathic Pain. Integrating Neurobiology, Neurodynamics and Clinical EvidenceDiego PaezAún no hay calificaciones
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeCalificación: 4 de 5 estrellas4/5 (5794)
- Anatomia e Fisiologia Da Lesão Do Nervo Sunderland1990Documento14 páginasAnatomia e Fisiologia Da Lesão Do Nervo Sunderland1990Ismael Vara CarvalhoAún no hay calificaciones
- Complementary and Alternative Medicine Treatments in PsychiatryDocumento111 páginasComplementary and Alternative Medicine Treatments in PsychiatryMaria IliescuAún no hay calificaciones
- The Little Book of Hygge: Danish Secrets to Happy LivingDe EverandThe Little Book of Hygge: Danish Secrets to Happy LivingCalificación: 3.5 de 5 estrellas3.5/5 (399)
- Avaliação Topográfica ASIDocumento7 páginasAvaliação Topográfica ASIIsmael Vara CarvalhoAún no hay calificaciones
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureCalificación: 4.5 de 5 estrellas4.5/5 (474)
- O Caminho Da Ciência - A Verdadeira HistóriaDocumento4 páginasO Caminho Da Ciência - A Verdadeira HistóriaIsmael Vara CarvalhoAún no hay calificaciones
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryCalificación: 3.5 de 5 estrellas3.5/5 (231)
- Complementary and Alternative Medicine Treatments in PsychiatryDocumento111 páginasComplementary and Alternative Medicine Treatments in PsychiatryMaria IliescuAún no hay calificaciones
- The Emperor of All Maladies: A Biography of CancerDe EverandThe Emperor of All Maladies: A Biography of CancerCalificación: 4.5 de 5 estrellas4.5/5 (271)
- Biomechanics vs. Pain Science - Bridging The Clinical Divide! (Part 1) - Accelerated Motion PhysiotherapyDocumento6 páginasBiomechanics vs. Pain Science - Bridging The Clinical Divide! (Part 1) - Accelerated Motion PhysiotherapyIsmael Vara CarvalhoAún no hay calificaciones
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersCalificación: 4.5 de 5 estrellas4.5/5 (344)
- Compensatory MechanismDocumento30 páginasCompensatory MechanismSenthil Kumar Elumalai100% (1)
- On Fire: The (Burning) Case for a Green New DealDe EverandOn Fire: The (Burning) Case for a Green New DealCalificación: 4 de 5 estrellas4/5 (73)
- Caso Clínico PieiraDocumento8 páginasCaso Clínico PieiraIsmael Vara CarvalhoAún no hay calificaciones
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaCalificación: 4.5 de 5 estrellas4.5/5 (265)
- Avaliação Entorses Tornozelo Com InstabilidadeDocumento8 páginasAvaliação Entorses Tornozelo Com InstabilidadeIsmael Vara CarvalhoAún no hay calificaciones
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyCalificación: 3.5 de 5 estrellas3.5/5 (2219)
- Heart and Circulatory SystemDocumento4 páginasHeart and Circulatory SystemIsmael Vara CarvalhoAún no hay calificaciones
- Team of Rivals: The Political Genius of Abraham LincolnDe EverandTeam of Rivals: The Political Genius of Abraham LincolnCalificación: 4.5 de 5 estrellas4.5/5 (234)
- Effects of Ibuprofen and Resistance Training On.2Documento8 páginasEffects of Ibuprofen and Resistance Training On.2Ismael Vara CarvalhoAún no hay calificaciones
- Kristiansen 2016Documento30 páginasKristiansen 2016Ismael Vara CarvalhoAún no hay calificaciones
- Tolerar A Incerteza A Próxima Revolução MédicaDocumento3 páginasTolerar A Incerteza A Próxima Revolução MédicaIsmael Vara CarvalhoAún no hay calificaciones
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreCalificación: 4 de 5 estrellas4/5 (1090)
- Dopamina e A AmigdalaDocumento6 páginasDopamina e A AmigdalaIsmael Vara CarvalhoAún no hay calificaciones
- Rise of ISIS: A Threat We Can't IgnoreDe EverandRise of ISIS: A Threat We Can't IgnoreCalificación: 3.5 de 5 estrellas3.5/5 (137)
- Biomechanics of The Osteoperotic SpineDocumento8 páginasBiomechanics of The Osteoperotic SpineIsmael Vara CarvalhoAún no hay calificaciones
- Dopamina e A AmigdalaDocumento6 páginasDopamina e A AmigdalaIsmael Vara CarvalhoAún no hay calificaciones
- The Unwinding: An Inner History of the New AmericaDe EverandThe Unwinding: An Inner History of the New AmericaCalificación: 4 de 5 estrellas4/5 (45)
- Dopamina e A AmigdalaDocumento6 páginasDopamina e A AmigdalaIsmael Vara CarvalhoAún no hay calificaciones
- Referências de Estudos Que Negam A Terapia ManualDocumento5 páginasReferências de Estudos Que Negam A Terapia ManualIsmael Vara CarvalhoAún no hay calificaciones
- Degeneração Dos Discos Implica Dor LombarDocumento10 páginasDegeneração Dos Discos Implica Dor LombarIsmael Vara CarvalhoAún no hay calificaciones
- Become A Clinical ExpertiseDocumento12 páginasBecome A Clinical ExpertiseIsmael Vara CarvalhoAún no hay calificaciones
- Efeito Da Manipulação Vertebral Nas Vias Motoras Do Córtex para Músculos Superiores e InferioresDocumento15 páginasEfeito Da Manipulação Vertebral Nas Vias Motoras Do Córtex para Músculos Superiores e InferioresIsmael Vara CarvalhoAún no hay calificaciones
- Dor Cervicobraquial Nem Sempre É NeurogénicaDocumento3 páginasDor Cervicobraquial Nem Sempre É NeurogénicaIsmael Vara CarvalhoAún no hay calificaciones
- Assessment and Treatment of Muscle Imbalance of Lower QuadrantDocumento87 páginasAssessment and Treatment of Muscle Imbalance of Lower QuadrantIsmael Vara CarvalhoAún no hay calificaciones
- Is Bacon Dangerous - The Science Behind Food Trends - Life and Style - The GuardianDocumento9 páginasIs Bacon Dangerous - The Science Behind Food Trends - Life and Style - The GuardianIsmael Vara CarvalhoAún no hay calificaciones
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Calificación: 4.5 de 5 estrellas4.5/5 (119)
- Elderly Suicide FactsDocumento2 páginasElderly Suicide FactsThe News-HeraldAún no hay calificaciones
- Sons and Lovers AuthorDocumento9 páginasSons and Lovers AuthorArmen NeziriAún no hay calificaciones
- Sample Essay: Qualities of A Good Neighbour 1Documento2 páginasSample Essay: Qualities of A Good Neighbour 1Simone Ng100% (1)
- SampleDocumento4 páginasSampleParrallathanAún no hay calificaciones
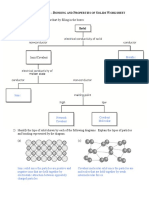
- Bonding and Properties of Solids Worksheet Solutions 1kadax6Documento4 páginasBonding and Properties of Solids Worksheet Solutions 1kadax6Mel Patricia M. CabreraAún no hay calificaciones
- The Perks of Being a WallflowerDe EverandThe Perks of Being a WallflowerCalificación: 4.5 de 5 estrellas4.5/5 (2099)
- Para Kay BDocumento1 páginaPara Kay BFeLy DipOn63% (8)
- International Journal of Current Advanced Research International Journal of Current Advanced ResearchDocumento4 páginasInternational Journal of Current Advanced Research International Journal of Current Advanced Researchsoumya mahantiAún no hay calificaciones
- Multiple Choice Test - 66253Documento2 páginasMultiple Choice Test - 66253mvjAún no hay calificaciones
- BRM 6Documento48 páginasBRM 6Tanu GuptaAún no hay calificaciones
- Predict Y Scores Using Linear Regression EquationDocumento6 páginasPredict Y Scores Using Linear Regression EquationreebenthomasAún no hay calificaciones
- Experiment 5 ADHAVANDocumento29 páginasExperiment 5 ADHAVANManoj Raj RajAún no hay calificaciones
- Her Body and Other Parties: StoriesDe EverandHer Body and Other Parties: StoriesCalificación: 4 de 5 estrellas4/5 (821)
- Dmat ReportDocumento130 páginasDmat ReportparasarawgiAún no hay calificaciones
- Us Aers Roadmap Noncontrolling Interest 2019 PDFDocumento194 páginasUs Aers Roadmap Noncontrolling Interest 2019 PDFUlii PntAún no hay calificaciones
- Engineering: Ronnapee Chaichaowarat, Jun Kinugawa, Kazuhiro KosugeDocumento8 páginasEngineering: Ronnapee Chaichaowarat, Jun Kinugawa, Kazuhiro Kosugelaura bemudezAún no hay calificaciones
- Audience AnalysisDocumento7 páginasAudience AnalysisSHAHKOT GRIDAún no hay calificaciones
- Detailed Lesson Plan in Science 10Documento7 páginasDetailed Lesson Plan in Science 10Glen MillarAún no hay calificaciones
- HexaflexDocumento10 páginasHexaflexCharlie Williams100% (1)
- Larry Dossey - HealingBeyondtheBodyDocumento2 páginasLarry Dossey - HealingBeyondtheBodypaulxeAún no hay calificaciones
- CV Jan 2015 SDocumento4 páginasCV Jan 2015 Sapi-276142935Aún no hay calificaciones
- Rhodes Solutions Ch4Documento19 páginasRhodes Solutions Ch4Joson Chai100% (4)
- One Shot To The HeadDocumento157 páginasOne Shot To The HeadEdison ChingAún no hay calificaciones
- Cost-Benefit Analysis of The ATM Automatic DepositDocumento14 páginasCost-Benefit Analysis of The ATM Automatic DepositBhanupriyaAún no hay calificaciones
- Analyzing Evidence of College Readiness: A Tri-Level Empirical & Conceptual FrameworkDocumento66 páginasAnalyzing Evidence of College Readiness: A Tri-Level Empirical & Conceptual FrameworkJinky RegonayAún no hay calificaciones
- Edwards 1999 Emotion DiscourseDocumento22 páginasEdwards 1999 Emotion DiscourseRebeca CenaAún no hay calificaciones
- 15Documento74 páginas15physicsdocs60% (25)
- Edukasyon Sa Pagpapakatao (Esp) Monitoring and Evaluation Tool For Department Heads/Chairmen/CoordinatorsDocumento3 páginasEdukasyon Sa Pagpapakatao (Esp) Monitoring and Evaluation Tool For Department Heads/Chairmen/CoordinatorsPrincis CianoAún no hay calificaciones
- Olimpiada Engleza 2017 CL A 7 A PDFDocumento4 páginasOlimpiada Engleza 2017 CL A 7 A PDFAnthony Adams100% (3)
- 317 Midterm QuizDocumento5 páginas317 Midterm QuizNikoruAún no hay calificaciones
- Cps InfographicDocumento1 páginaCps Infographicapi-665846419Aún no hay calificaciones
- ME Flowchart 2014 2015Documento2 páginasME Flowchart 2014 2015Mario ManciaAún no hay calificaciones
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionDe EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionCalificación: 4 de 5 estrellas4/5 (402)
- Why We Die: The New Science of Aging and the Quest for ImmortalityDe EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityCalificación: 3.5 de 5 estrellas3.5/5 (2)
- The 7 Habits of Highly Effective People: The Infographics EditionDe EverandThe 7 Habits of Highly Effective People: The Infographics EditionCalificación: 4 de 5 estrellas4/5 (2475)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsDe EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsCalificación: 3.5 de 5 estrellas3.5/5 (3)
- The Stoic Mindset: Living the Ten Principles of StoicismDe EverandThe Stoic Mindset: Living the Ten Principles of StoicismAún no hay calificaciones
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeDe EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeAún no hay calificaciones
- Summary: Atomic Habits by James Clear: An Easy & Proven Way to Build Good Habits & Break Bad OnesDe EverandSummary: Atomic Habits by James Clear: An Easy & Proven Way to Build Good Habits & Break Bad OnesCalificación: 5 de 5 estrellas5/5 (1631)
- The Power of Now: A Guide to Spiritual EnlightenmentDe EverandThe Power of Now: A Guide to Spiritual EnlightenmentCalificación: 4.5 de 5 estrellas4.5/5 (4120)